
Prevalence of Biofilm Formation and Quorum Sensing Genes in Pseudomonas aeruginosa and Escherichia coli Isolated from Smoked Fish
 , Moumouny Traore2 , Benie Comoe Koffi Donatien1 , Atobla Koua1 and Dadie Adjehi3
, Moumouny Traore2 , Benie Comoe Koffi Donatien1 , Atobla Koua1 and Dadie Adjehi3 1Laboratory of Biotechnology Agriculture and Valorization of Biological Resource, UFR Biosciences, Felix Houphouet Boigny University, Abidjan, Côte d’Ivoire
2Laboratory of Food Microbiology and Biotechnology, Department of Biochemistry and Genetics, Peleforo Gon Coulibaly University, Korhogo, Côte d’Ivoire
3Laboratory of Food Microbiology and Biotechnology, Department of Food Science and Technology, Nangui Abrogoua University, Abidjan, Côte d'Ivoire
Corresponding Author E-mail: k_nzebo@yahoo.fr
DOI : http://dx.doi.org/10.13005/bbra/3533
Download this article as:
![]()
This investigation sought to ascertain the ability of Pseudomonas aeruginosa and Escherichia coli isolated from contaminated smoked fish to form biofilms and engage in quorum sensing. Sixty strains of Pseudomonas aeruginosa and forty strains of Escherichia coli isolated from smoked fish were used to conducted this study. The strains were first revived and then inoculated onto Cetrimide and Tryptone bile X glucuronide media. Quorum sensing (LasI and LasR) and biofilm formation gene such as ppyR, PelA and PslA were detected by PCR methods. In P. aeruginosa strains, genes Las were detected with a prevalence of 75% for LasI and LasR. The prevalence of Rhl genes was 58.33% for RhlI and 50% for RhlR, respectively. In E. coli strains, the prevalence was 71.42% for the genes (RhlI/R) and 42.85% for the genes (LasI/R). The biofilm-forming genes PslA, ppyR and PelA were detected in Pseudomonas aeruginosa strains with respective percentages of 50%, 41.66%, and 58.33%. Only PelA (14.28%) and PslA (28.57%) were detected in Escherichia coli strains. Pseudomonas aeruginosa and Escherichia coli are involved in biofilm formation and quorum sensing. Smoking techniques and smoked fish preservation methods need to be improved.
KEYWORDS:Biofilm; Escherichia coli; Pseudomonas aeruginosa; Quorum sensing; Smoked fish
Introduction
Biofilms are complex and interconnected bacterial communities. They can adhere to various surfaces in the food industry, kitchens (cutting boards, plumbing fixtures), and even on equipment used in hospitals.1 Extracellular polymeric substances (EPS), proteins, lipids, and extracellular DNA are the most important components in the formation of the biofilm’s complex matrix. Biofilm shielded bacteria from antimicrobial agents, host immunological responses, and adverse surroundings.2-4 Biofilm formation is considered a bacterial survival strategy in hostile environments.5 A matrix of extracellular polymeric materials envelops and shields the bacterial communities residing in biofilms. The bacterial species and ambient conditions determine the physicochemical characteristics and makeup of these extracellular polymeric materials.6
Food contamination and spoiling can result from pathogenic bacteria like Escherichia coli and Pseudomonas aeruginosa growing and sticking to the surfaces of tools and utensils. These bacteria are commonly found in fish. Their participation in adhesion processes is well-known, biofilm formation, and quorum sensing in humans and animals.3,7 Furthermore, they are implicated in nosocomial infections and food poisoning through a diversity of virulence factors mediated by quorum sensing (QS).8,9
Biofilms have a major impact on the pathogenicity and antibiotic resistance of Pseudomonas aeruginosa. Pseudomonas aeruginosa needs type IV pili and a single polar flagellum in order to attach, colonize surfaces, and form biofilms. He can survive on biotic and abiotic surfaces while eluding human immune responses thanks to biofilms.10 Pseudomonas aeruginosa associated with biofilms exhibits increased antibiotic resistance.11-13
Escherichia coli is not naturally present in the fish microbiota but can be introduced from contaminated aquatic environments or during fish handling.14 The presence of Escherichia coli in food, including fish, is considered an indicator of faecal contamination.15 Although most Escherichia coli strains are non-pathogenic, various intestinal serotypes are opportunistic pathogens and can cause gastrointestinal, urinary, or central nervous system diseases.16 Escherichia coli possesses both the ability to form biofilms and antibiotic resistance.17,18 Consequently, they can survive for many years in food-processing environments.19
The increase in antimicrobial resistance among food-producing animals, including seafood, may aid in the food chain’s spread to humans.20,21 Certain pathogens’ capacity to build biofilms makes treatment management even more difficult. Biofilms act as physical barriers, significantly reducing the effectiveness of antibiotics and immune recognition.22 The presence of pathogenic bacteria in biofilms that form on the contact surfaces of fumigation ovens and display counters poses a potential risk of disease transmission to consumers.23
Acting as physical barriers, biofilms markedly diminish the efficacy of antibiotics and impair immune recognition.22 When pathogenic bacteria are embedded within biofilms on food-contact surfaces (such as smoking ovens or fish display counters) they can serve as persistent reservoirs of contamination and contribute to human infections.23
Biofilms formed from hospital and animal strains have been studied in Côte d’Ivoire.24 However, Research on quorum sensing and biofilm formation in smoked fish is virtually non-existent. The overarching objective of this study was to determine the genetic potential for quorum sensing and biofilm formation in Escherichia coli and Pseudomonas aeruginosa isolated from smoked fish sold in markets across Abidjan.
Materials and Methods
Materials
Biological Material
The biological material consisted of multidrug-resistant strains of Pseudomonas aeruginosa and Escherichia coli, isolated from smoked fish. These strains originated from a previous study.25
Methods
Confirmation of strains
Colonies characteristic of Pseudomonas aeruginosa and Escherichia coli were streaked onto selective and non-selective agar plates. Gram staining, the fresh-state test, and the classic LEMINOR miniature gallery test were performed on presumptive Escherichia coli colonies after 18 to 24 hours of incubation at 37°C.
After 24 hours of growth on Luria-Bertani Agar (LB), the oxidase-positive strains were cultured on King A and King B media. Those producing both types of pigment: pyoverdin (yellow-green, fluorescent pigment) on King B and pyocyanin (blue, non-fluorescent pigment) on King A, or pyocyanin alone on King A, and capable of growing at 42°C were identified as Pseudomonas aeruginosa. Oxidase-positive strains that did not produce pyocyanin and could not grow at 42°C were considered Pseudomonas sp.
Preservation of isolates
The identified bacterial strains of Pseudomonas aeruginosa and Escherichia coli was sub-cultured in duplicate into Tryptic Soy Broth (TSB) (Sigma-Aldrich)-filled hemolysis tubes, followed add 500 µL of glycerol to reach a final concentration of 15%. After 3 hours of growth at 28 °C, the cultures were preserved transiently for 1 hour at –20 °C and subsequently stored at –80 °C.
Identification of genes implicated in the development of biofilms
Amplification of genes: ppyR, pslA, pelA
The technique outlined by Pournajaf et al.26 was used to amplify the biofilm formation genes (pslA, pelA, and ppyR).26 The amplification program comprised a repeated cyclic phase after an initial denaturation for five minutes at 95 °C. A denaturation step of 30 seconds at 95°C, a primer annealing step of 60 seconds at 65°C, and an elongation step of 90 seconds at 72°C made up this cyclic phase, which was repeated 33 times. A final elongation of five minutes at 72°C marked the end of the amplification reaction. On a T9700 Thermocycler (Applied Biosystems, China), samples were kept at +4°C until the run was finished. The nucleotide sequences of the primers used are presented in Table 1.
Amplification of quorum sensing genes (LasI, LasR, rhlI, rhlR)
Quorum sensing genes (LasI, LasR, rhlI, and rhlR) were amplified using the procedure outlined by Lima et al.27 The amplification program comprised a cyclic phase that was repeated 33 times after an initial denaturation for five minutes at 95 °C. Each cycle included a 30-second denaturation step at 95°C, a 60-second primer annealing stage at 65°C, and a 90-second elongation step at 72°C. A final elongation of five minutes at 72°C marked the end of the amplification reaction. Samples were kept on a Thermocycler (Applied Biosystems) at +4°C until the run was finished. The nucleotide sequences of the primers used are presented in Table 1.
Electrophoresis of amplification products
A transilluminator was used to view the bands under UV light after they had migrated at 120 V for 30 minutes on an 8% agarose gel. A Molecular Imager Gel DocTM EZ system (Bio-Rad, USA) was then used to take pictures of them.
Table I: Primers used for the amplification of ppyR, pslA, pelA genes and LasI, LasR, rhlI, rhlR
| Genes | Primers sequences (5ʹ→3ʹ) | Base pair size | |
| Biofilm | pslA
|
F : 5’-TCCCTACCTCAGCAGCAAGC-3’
R : 5’-TGTTGTAGCCGTAGCGTTTCTG-3’ |
656 |
| PelA | F : 5’-CATACCTTCAGCCATCCGTTCTTC-3’
R : 5’-CGCATTCGCCGCACTCAG-3’ |
786 | |
| PpyR | F : 5’-CGTGATCGCCGCCTATTTCC-3’
R : 5’-ACAGCAGACCTCCCAACCG-3’ |
160 | |
| quorum sensing | LasI
|
F : 5’-CGTGCTCAAGTGTTCAAGG -3’
R : 5’-TACAGTCGGAAAAGCCCAG -3’ |
295 |
| LasR
|
F : 5’-AAGTGGAAAATTGGAGTGGAG-3’
R : 5’-GTAGTTGCCGACGACGATGAAG- 3’ |
130 | |
| rhlI
|
F : 5’-TTCATCCTCCTTTAGTCTTCCC-3’
R : 5’-TTCCAGCGATTCAGAGAGC -3’ |
155 | |
| rhlR | F : 5’-TGCATTTTATCGATCAGGGC-3’
R : 5’-CACTTCCTTTTCCAGGACG-3’ |
133 | |
| IMP | F : TGAGCAAGTTATCTGTATTC
R : TTAGTTGCTTGGTTTTGATG |
740 |
Calculation of the prevalence of isolated genes.
To determine the prevalence of individual genes, the following formula was used.
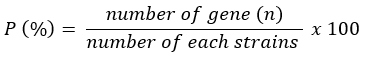
Data analysis
To investigate the possible relationship between the genes involved in biofilm formation and quorum sensing and the isolated bacterial strains, a principal component analysis (PCA) was performed.
Results
Quorum sensing and biofilm-associated genes detected by PCR
Molecular determinism of quorum sensing genes: LasI, LasR, rhlI, rhlR
Figure 1 illustrates the electrophoretic profiles of genetic determinants involved in quorum sensing in multidrug-resistant Escherichia coli and Pseudomonas aeruginosa. Certain strains of Escherichia coli and Pseudomonas aeruginosa harbored the Las or Rhl genes, which encode elastase and rhamnolipid production, respectively. In Pseudomonas aeruginosa, the prevalence of Las genes was 77.41% for LasI and 77.41% for LasR. The prevalence of Rhl genes was 58.06% for RhlI and 54.83% for RhlR. In E. coli, Las and Rhl genes were detected with respective prevalences of 38.88% (LasI/R) and 72.22% (RhlI/RhlR) (Table 2).
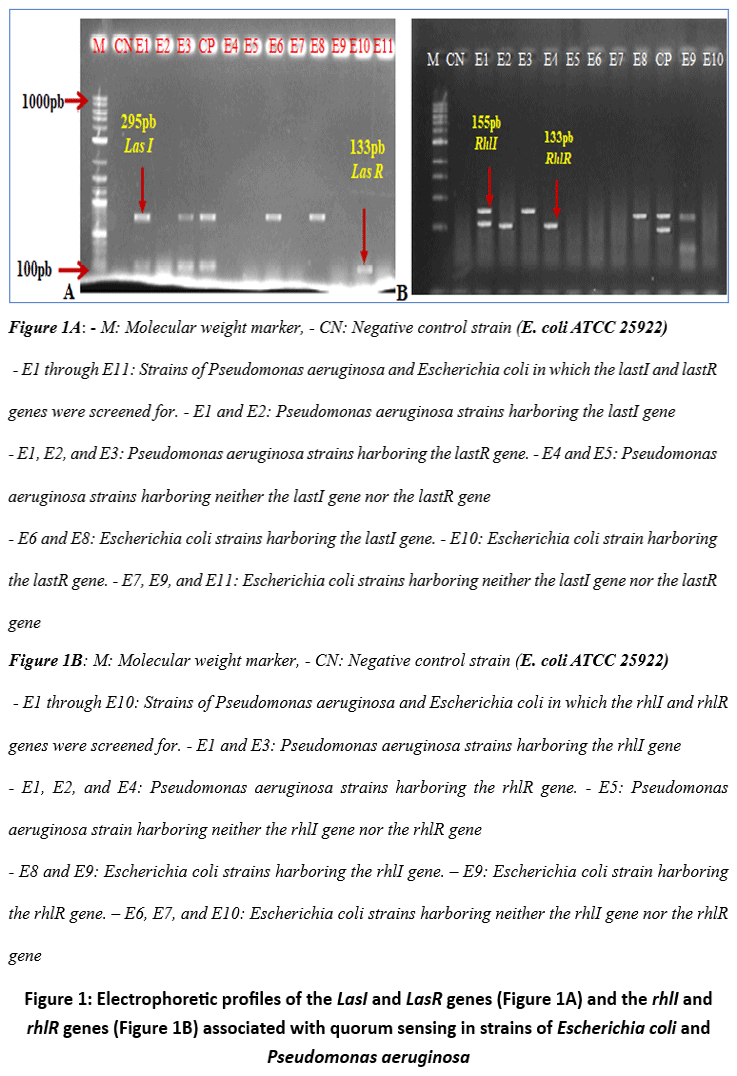 |
Figure 1: Electrophoretic profiles of the LasI and LasR genes (Figure 1A) and the rhlI and rhlR genes (Figure 1B) associated with quorum sensing in strains of Escherichia coli and Pseudomonas aeruginosa.
|
Table 2: Prevalence of genes associated with quorum sensing in Pseudomonas aeruginosa and Escherichia coli
| Quorum sensing genes | P. aeruginosa (n=12) | E. coli (n=7) | Total (n=19) |
| Prevalence (%) and number of cases | |||
| LasI | 75,0 (9) | 42,85 (3) | 63,15 (12) |
| LasR | 75,0 (9) | 42,85 (3) | 63,15 (12) |
| RhlI | 58,33 (7) | 71,42 (5) | 63,15 (12) |
| RhlR | 50,0 (6) | 71,42 (5) | 57,89 (11) |
Las gene: Encodes the production of elastase; Rhl gene: Encodes the production of rhamnolipid
Molecular determinants of biofilm: ppyR, PelA and PslA
Figure 2 shows the electrophoretic profiles of biofilm‑forming genes in multidrug‑resistant strains of Pseudomonas aeruginosa and Escherichia coli. All analyzed P. aeruginosa strains carried biofilm‑associated genes. The three biofilm‑forming genes (ppyR, PelA and PslA) were detected in some Pseudomonas strains, while others possessed two or only one of these genes. The biofilm‑forming genes PslA, PelA, and ppyR were detected in all twelve multidrug‑resistant Pseudomonas aeruginosa strains (Figure 2A). In the case of Escherichia coli strains, some carried the biofilm‑associated genes PslA and/or PelA, whereas others did not present any of them (Figure 2B).
Prevalences of PslA, PelA, and ppyR were respectively 50.0%, 41.66%, and 58.33%. In contrast, among the seven multidrug‑resistant Escherichia coli strains, only the biofilm‑forming genes PslA and PelA were identified, with prevalences of 28.57% and 14.28%, respectively. No gene encoding pyoverdine production (ppyR) was detected in E. coli (Table 3).
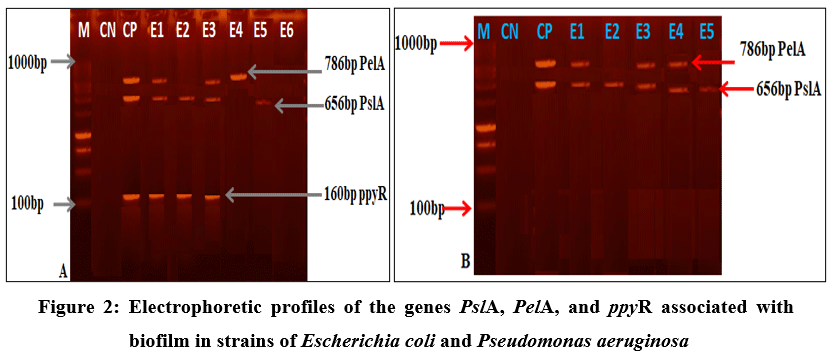 |
Figure 2: Electrophoretic profiles of the genes PslA, PelA, and ppyR associated with biofilm in strains of Escherichia coli and Pseudomonas aeruginosa
|
Table 3: Prevalence of genes associated with Biofilm in Pseudomonas aeruginosa and Escherichia coli
| P. aeruginosa (n=12) | E. coli (n=7) | |
| Biofilms-associated genes | Prevalence (%) and number of cases | |
| PslA | 50,0% (6) | 28,57% (2) |
| PelA | 41,66 % (5) | 14,28% (1) |
| PpyR | 58,33% (7) | 0,0% (0) |
| PslA-PelA | 16,66% (2) | 33,33% (5) |
| PslA-PpyR | 8,33% (1) | 0% (0) |
| PelA- PpyR | 0% (0) | 0% (0) |
| PslA-PelA-PpyR | 25,0% (3) | 0% (0) |
Pel: pellicle (gene encoding pellicle formation); Psl: polysaccharide synthesis locus (polysaccharide synthesis); ppyR: psl and pyoverdine operon regulator, PA2663 (regulator of the psl and pyoverdine operons)
Distribution of biofilm‑forming genes in Pseudomonas aeruginosa and Escherichia coli
Principal component analysis (PCA) axes 1 and 2 together account for 100% of the total variance. Axis 1 of the PCA individual plot shows that the biofilm‑forming genes PslA and PelA are more closely associated with Escherichia coli strains than with Pseudomonas aeruginosa strains, which are the only ones harboring ppyR. However, axis 2 indicates that P. aeruginosa strains are more strongly associated with the genes ppyR and PslA. (Figure 3).
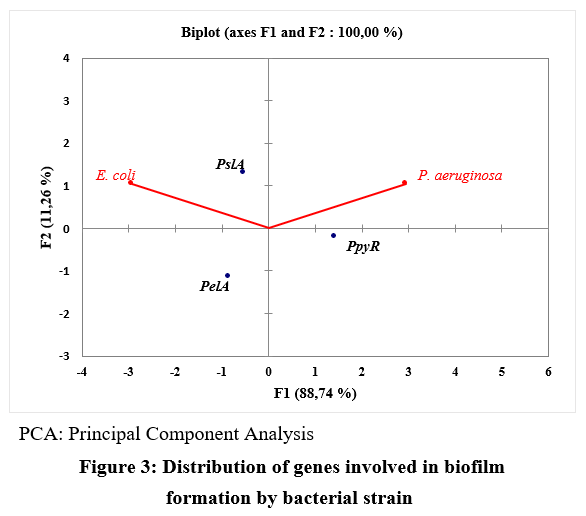 |
Figure 3: Distribution of genes involved in biofilm formation by bacterial strain
|
Discussion
In this investigation, isolates of Escherichia coli and Pseudomonas aeruginosa were found to contain genetic components involved in quorum sensing and biofilm development. The Las or Rhl genes were present in certain strains of Escherichia coli and Pseudomonas aeruginosa. The Las gene encodes elastase production, whereas the Rhl gene encodes rhamnolipid production. These genes were detected in P. aeruginosa isolates with prevalences of 75.0% for LasI/R, 58.33% for RhlI, and 50.0% for RhlR. In E. coli, Las and Rhl genes were detected with prevalences of 42.85% (LasI/R) and 71.42% (RhlI/R), respectively. The relatively high frequencies of these autoinducers in P. aeruginosa and E. coli strains may support the existence of a communication system between these bacteria isolated from animal products. Quorum sensing in these strains of P. aeruginosa and E. coli may be under the control of the LasI/R and RhlI/R systems, which are involved in the synthesis of certain virulence genes.28,29
In Gram‑negative bacteria, two main QS effector/receptor systems involving homoserine lactones coexist and regulate the expression of virulence genes.30 The 3‑oxo‑C12‑homoserine lactone are involved in the LasI/R system, which codes for virulence factors like elastases LasA and LasB and exotoxin A (ToxA). The second system, RhlI/R, is responsible for the production and regulation of factors such as rhamnolipids, pyocyanin, and cyanide, and involves C4‑homoserine lactone (C4‑HSL).31,32 The presence of Las and Rhl genes in both Pseudomonas aeruginosa and Escherichia coli isolates from smoked fish suggests that they may be involved in the production of bacterial pheromones that facilitate quorum sensing.
Nutrient limitation, stress, and cell density influence the biosynthesis of rhamnolipids.33,34 In addition to the genes involved in quorum sensing, this study identified several genes encoding biofilm formation. Specifically, genes ppyR, pslA, and pelA associated to the biofilm were detected in Pseudomonas aeruginosa with respective prevalences of 50.0%, 41.66%, and 58.33%. The principal genes ppyR, pslA and pelA detected in P. aeruginosa strains suggest that these isolates are respectively implicated in the synthesis of pyoverdine, polysaccharide, and pellicle.
The production of pyoverdine, polysaccharide, and pellicle observed in these strains could promote mechanisms involved in the pathogenicity of P. aeruginosa through quorum sensing.32,35 In Escherichia coli, the same genes pelA (14.28%) and pslA (28.57%), implicated in the biosynthesis of polysaccharide and pellicle, were detected; however, no gene encoding pyoverdine production was identified. The absence of the ppyR gene in E. coli accounts for the lack of pyoverdine production in this strain.
Principal component analysis (PCA) confirmed that the biofilm‑forming genes pslA and pelA are more closely associated with Escherichia coli strains than with Pseudomonas aeruginosa strains, which are the only isolates harboring ppyR.
The presence of numerous resistance genes, together with evidence of quorum sensing and biofilm formation, underscores the need for reliable and appropriate food preservation procedures and techniques, particularly for products of animal origin, throughout the processing chain. Their occurrence in food, especially smoked fish, represents a major public health, and their epidemic potential.
Conclusion
This study demonstrated that Pseudomonas aeruginosa and Escherichia coli isolated from smoked fish sold in markets in Abidjan harbored quorum sensing and biofilm formation gene. Quorum sensing genes (las, rhl) and biofilm‑associated genes (pslA, pelA, ppyR) were identified in the isolates. No ppyR gene was detected in E. coli. The study also showed that all of these determinants may contribute to the virulence and pathogenicity of strains isolated from smoked fish. Improvements in smoking and preservation techniques for smoked fish are therefore required. Microorganisms found in smoked fish and the consumption of these products must be monitored. It is therefore recommended that vendors of smoked fish improve their work environment and adhere to hygiene standards regarding the workplace, equipment, and handlers, as well as proper packaging techniques for smoked fish. The presence of genes involved in biofilm formation and quorum sensing should serve as a warning to prevent public health issues.
Acknowledgement
The authors acknowledge the Biotechnology Laboratory of the Department of Biosciences at Felix Boigny University for Characterization studies.
Funding Sources
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Conflict of Interest
The authors do not have any conflict of interest.
Data Availability Statement
This statement does not apply to this article.
Ethics Statement
This research did not involve human participants, animal subjects, or any material that requires ethical approval.
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Clinical Trial Registration
This research does not involve any clinical trials.
Permission to reproduce material from other sources
Not Applicable.
Author Contributions
- N’zebo Desire KOUAME: Conceptualization, Validation, Methodology, Writing – original draft, Writing – review & editing. Visualization, Formal analysis.
- Moumouny TRAORE: Analysis and interpretation of data, – writing the article.
- Benie Comoe Koffi Donatien: Investigation, Formal analysis.
- Atobla Koua: Investigation, – proofreading and editing the manuscript;
- Dadie Adjehi: Formal analysis, – final approval of the version to be published
References
- Xu Z, Liu Z, Soteyome T, et al. Impact of pmrA on Cronobacter sakazakii planktonic and biofilm cells: A comprehensive transcriptomic study. Food Microbiol. 2021;(98):103785. doi: 10.1016/j.fm.2021.103785.
CrossRef - Yuan L, Sadiq FA, Wang N, Yang Z, He G. Recent advances in understanding the control of disinfectant-resistant biofilms by hurdle technology in the food industry. Crit Rev Food Sci Nutr. 2021;61(22):3876-3891. doi: 10.1080/10408398.2020.1809345.
CrossRef - Li X, Gu N, Huang TY, Zhong F, Peng G. Pseudomonas aeruginosa: A typical biofilm forming pathogen and an emerging but underestimated pathogen in food processing. Front Microbiol. 2023;(13):1114199. doi: 10.3389/fmicb.2022.1114199.
CrossRef - Wen S, Mai Y, Chen X, et al. Molecular epidemiology and antibiotic resistance analysis of non-typeable Haemophilus influenzae (NTHi) in Guangzhou: A representative city of southern China. Antibiotics. 2023;12(4):656. doi: 10.3390/antibiotics12040656.
CrossRef - Moradali MF, Ghods S, Rehm BH. Pseudomonas aeruginosa Lifestyle: A paradigm for adaptation, survival, and persistence. Front Cell Infect Microbiol. 2017;(7):39. doi: 10.3389/fcimb.2017.00039.
CrossRef - Flemming HC, Wingender J. The biofilm matrix. Nat Rev Microbiol. 2010;8(9):623-33. doi: 10.1038/nrmicro2415.
CrossRef - Mataragka A, Anthi R, Christodouli ZE, Malisova O, Andritsos ND. Presence of major bacterial foodborne pathogens in the domestic environment and hygienic status of food cleaning utensils: A narrative review. Hygiene. 2025;5(4):60. org/10.3390/hygiene5040060
CrossRef - Streeter K, Katouli M. Pseudomonas aeruginosa: A review of their pathogenesis and prevalence in clinical settings and the environment. epidemiol. med. 2016;2(1):25–32. doi.org/10.18869/modares.iem.2.1.25
CrossRef - Virupakshaiah DBM, Hemalata VB. Molecular identification of Pseudomonas aeruginosa from food borne isolates. J. Curr. Microbiol. App. Sci. 2016;5(6):1026–1032. doi.org/10.20546/ijcmas.2016.506.109
CrossRef - De Sousa T, Hébraud M, Dapkevicius MLNE, et al. Genomic and metabolic characteristics of the pathogenicity in Pseudomonas aeruginosa. Int J Mol Sci. 2021;22(23):12892. doi: 10.3390/ijms222312892.
CrossRef - Quintieri L, Fanelli F, Caputo L. Antibiotic resistant Pseudomonas spoilers in fresh dairy products: An underestimated risk and the control strategies. Foods. 2019;8(9):372. doi.org/10.3390/foods8090372
CrossRef - Sharma S, Mohler J, Mahajan SD, et al. Microbial Biofilm: A Review on Formation, Infection, Antibiotic Resistance, Control Measures, and Innovative Treatment. Microorganisms 2023, 11, 1614. Microorganisms. 2024;12(10):1961. doi: 10.3390/microorganisms12101961.
CrossRef - Liang J, Huang TY, Li X, Gao Y. Germicidal effect of intense pulsed light on Pseudomonas aeruginosa in food processing. Front Microbiol. 2023 ;(14):1247364. doi: 10.3389/fmicb.2023.1247364.
CrossRef - Cardozo MV, Borges CA, Beraldo LG, et al. Shigatoxigenic and atypical enteropathogenic Escherichia coli in fish for human consumption. Braz J Microbiol. 2018;49(4):936–941. org/10.1016/j.bjm.2018.02.013
CrossRef - Prakasan S, Lekshmi M, Ammini P, Balange AK, Nayak BB, Kumar SH. Occurrence, pathogroup distribution and virulence genotypes of Escherichia coli from fresh seafood. Food Control. 2022;(133):108669.
CrossRef - Fadel A, Mabrok M, Aly S. Epizootics of Pseudomonas anguilliseptica among cultured seabream (Sparus aurata) populations: Control and treatment strategies. Microb Pathog. 2018;(121):1-8. doi: 10.1016/j.micpath.2018.04.021.
CrossRef - Shrestha A, Chaudhary CK, Ghale R, Shrestha, A. Monosex male tilapia production. Int J Anim Biotechnol Appl. 2019;4(2):18-23
- Qian W, Li X, Yang M, et al. Relationship between antibiotic resistance, biofilm formation, and biofilm-specific resistance in Escherichia coli Isolates from Ningbo, China. Infect Drug Resist. 2022;(15):2865-2878. doi: 10.2147/IDR.S363652.
CrossRef - Trang PN, Anh Ngoc TT, Masuda Y, Hohjoh KI, Miyamoto T. Antimicrobial resistance and biofilm formation of Escherichia coli in a Vietnamese Pangasius fish processing facility. Heliyon. 2023;9(10):e20727. doi: 10.1016/j.heliyon.2023.e20727.
CrossRef - Saharan VV, Verma P, Singh AP. High prevalence of antimicrobial resistance in Escherichia coli, Salmonella and Staphylococcus aureus isolated from fish samples in India. Aquac. Res. 2019;51(3);1200–1210. doi.org/10.1111/are.14471
CrossRef - Mwanza F, Komba EVG, Kambarage DM. Occurrence and determination of antimicrobial resistant Escherichia coli isolates in fish and vegetables as indicator organism of faecal contamination in Dar es Salaam, Tanzania. Int J Microbiol. 2021;(2021):6633488. doi: 10.1155/2021/6633488.
CrossRef - Thi MTT, Wibowo D, Rehm BHA. Pseudomonas aeruginosa Int J Mol Sci. 2020;21(22):8671. doi: 10.3390/ijms21228671.
CrossRef - Sofos JN, Geornaras I. Overview of current meat hygiene and safety risks and summary of recent studies on biofilms, and control of Escherichia coli O157:H7 in nonintact, and Listeria monocytogenes in ready-to-eat, meat products. Meat Sci. 2010;86(1):2-14. doi: 10.1016/j.meatsci.2010.04.015.
CrossRef - Benie CKD, Attien YP, Atobla K, Tra BYC, Dosso M. Quorum sensing regulated by N-acyl-homoserine lactone production and inhibitory effect of essential oils in aeruginosa. J Bacteriol Mycol. 2021;(9):4-20
- Azon CN. Antibiorésistance et biofilms de Pseudomonas aeruginosa, coli et Staphylococcus aureus contaminants les poissons fumes en Côte d’Ivoire. Master Microbiologie et Biotechnologie des aliments. Université Nangui Abrogoua. 2022 ; pp 54.
- Pournajaf A, Rajabnia R, Razavi S, et al. Molecular characterization of carbapenem-resistant; Acinetobacter baumannii; isolated from pediatric burns patients in an Iranian hospital. 2018;17(1):135-142. https://doi.org/10.4314/tjpr.v17i1.19
CrossRef - Lima JLDC, Alves LR, Jacomé PRLA, Bezerra Neto JP, Maciel MAV, Morais MMC. Biofilm production by clinical isolates of Pseudomonas aeruginosa and structural changes in LasR protein of isolates non-biofilm-producing. Braz J Infect Dis. 2018;22(2):129-136. doi: 10.1016/j.bjid.2018.03.003.
CrossRef - Morohoshi T, Fukamachi K, Kato M, Kato N, Ikeda T. Regulation of the violacein biosynthetic gene cluster by acylhomoserine lactone-mediated quorum sensing in Chromobacterium violaceum ATCC 12472. Biosci Biotechnol Biochem. 2010;74(10):2116-9. doi: 10.1271/bbb.100385
CrossRef - Zhu H, He CC, Chu QH. Inhibition of quorum sensing in Chromobacterium violaceum by pigments extracted from Auricularia auricular. Lett Appl Microbiol. 2011;52(3):269-274. doi: 10.1111/j.1472-765X.2010.02993.x.
CrossRef - Rasamiravaka T, Labtani Q, Duez P, El Jaziri M. The formation of biofilms by Pseudomonas aeruginosa: a review of the natural and synthetic compounds interfering with control mechanisms. Biomed Res Int. 2015;(2015):759348. doi: 10.1155/2015/759348.
CrossRef - Simanek K A, Schumacher M L, Mallery C P, Shen S, Li L, Paczkowski J E. Quorum-sensing synthase mutations re-calibrate autoinducer concentrations in clinical isolates of Pseudomonas aeruginosa to enhance pathogenesis. Commun. 2023;14(1)7986. doi.org/10.1038/s41467-023-43702-4
CrossRef - Maisuria VB, Los Santos YL, Tufenkji N, Déziel E. Cranberry-derived proanthocyanidins impair virulence and inhibit quorum sensing of Pseudomonas aeruginosa. Sci Rep. 2016;6(1):30169. doi: 10.1038/srep30169.
CrossRef - Dekimpe V, Déziel E. Revisiting the quorum-sensing hierarchy in Pseudomonas aeruginosa: the transcriptional regulator RhlR regulates LasR-specific factors. Microbiology. 2009;155(3):712-723. doi: 10.1099/mic.0.022764-0.
CrossRef - Déziel E, Lépine F, Milot S, He J, Mindrinos MN, Tompkins RG, Rahme LG. Analysis of Pseudomonas aeruginosa 4-hydroxy-2-alkylquinolines (HAQs) reveals a role for 4-hydroxy-2-heptylquinoline in cell-to-cell communication. Proc Natl Acad Sci U S A. 2004;101(5):1339-44. doi: 10.1073/pnas.0307694100.
CrossRef - Nagata T, Oobo T, Aozasa O. Efficacy of a bacterial siderophore, pyoverdine, to supply iron to Solanum lycopersicum J Biosci Bioeng. 2013;115(6):686-90. doi: 10.1016/j.jbiosc.2012.12.018
CrossRef
Accepted on: 02-06-2026
Second Review by: Dr. Makhabbah Jamilatun
Final Approval by: Dr. Jagdish Chandra Joshi








![]()
![]()
