
Neurophytopharmacology in Depression: Emerging Antidepressant Potential of Medicinal Plants from Experimental Animal Models
 , Kishor Otariand Ajay Kale
, Kishor Otariand Ajay Kale Department of Pharmacology, Navsahyadri Institute of Pharmacy, Naigaon, Pune, India.
Corresponding Author E-mail: damayantik2002@gmail.com
DOI : http://dx.doi.org/10.13005/bbra/3519
Download this article as:
![]()
Depression is a highly prevalent neuropsychiatric disorder characterized by persistent low mood, anhedonia, cognitive dysfunction, and impaired quality of life. Despite the availability of pharmacological treatments, their therapeutic outcomes remain suboptimal due to delayed onset of action, incomplete remission, and significant adverse effects. These limitations have prompted increasing interest in alternative therapeutic approaches, particularly those derived from medicinal plants. Neurophytopharmacology, an emerging interdisciplinary field, focuses on the interaction between plant-derived bioactive compounds and the central nervous system. Experimental evidence from animal models, such as the forced swim test, tail suspension test, and chronic unpredictable mild stress model, has demonstrated significant antidepressant-like activity of various plant extracts. Phytochemicals, including flavonoids, alkaloids, terpenoids, and phenolic compounds, exert their effects through multiple mechanisms, including modulation of monoaminergic neurotransmission, attenuation of oxidative stress, suppression of neuroinflammation, and enhancement of neuroplasticity. This review summarizes the current experimental evidence on medicinal plants with antidepressant potential, their mechanisms of action, and their relevance in preclinical models of depression.
KEYWORDS:Animal models; Antidepressant activity; Depression; Herbal medicine; Medicinal plants; Neurophytopharmacology; Neuroprotection; Phytochemicals
Introduction
Depression is a complex and multifactorial neuropsychiatric disorder that profoundly affects emotional well-being, cognitive function, and overall quality of life.1,2 It is clinically characterized by persistent low mood, loss of interest or pleasure in daily activities (anhedonia), fatigue, sleep disturbances, impaired concentration, and feelings of hopelessness or worthlessness.3 These symptoms often lead to significant impairment in social, occupational, and personal functioning, thereby reducing an individual’s ability to perform routine activities effectively.4 Depression is recognized as one of the most prevalent mental health disorders worldwide and represents a major public health concern.1 The global burden associated with depressive disorders continues to rise, affecting individuals across different age groups, socioeconomic backgrounds, and geographic regions.4,1
The pathogenesis of depression is highly complex and involves the interaction of multiple biological, psychological, and environmental factors.3 Several neurobiological mechanisms have been proposed to explain the development and progression of depressive disorders. Among these, the monoamine hypothesis has historically served as a central framework, suggesting that alterations in neurotransmitters such as serotonin, norepinephrine, and dopamine play a critical role in the manifestation of depressive symptoms.5,6 However, growing scientific evidence indicates that depression cannot be fully explained by neurotransmitter imbalance alone, highlighting the involvement of additional biological pathways.6
Recent research has increasingly focused on mechanisms such as neuroinflammation, oxidative stress, mitochondrial dysfunction, and impaired neuroplasticity.7,9 Chronic activation of inflammatory pathways and excessive production of reactive oxygen species can result in neuronal damage and functional impairment in key brain regions involved in mood regulation.8,9 These pathological changes may disrupt neuronal communication and contribute to the persistence of depressive symptoms. Furthermore, dysregulation of the hypothalamic–pituitary–adrenal (HPA) axis, often triggered by prolonged exposure to stress, has been strongly implicated in depression.10,11 Sustained elevation of glucocorticoids, particularly cortisol, can negatively affect neuronal survival and lead to structural and functional alterations in the hippocampus, prefrontal cortex, and amygdala.4,5
Pharmacological management of depression primarily involves the use of antidepressant medications such as selective serotonin reuptake inhibitors, tricyclic antidepressants, monoamine oxidase inhibitors, and other newer drug classes.2 Although these therapies have significantly improved clinical management, their therapeutic effectiveness is often limited by the delayed onset of action, incomplete remission of symptoms, and high rates of relapse.23 Additionally, conventional antidepressants are frequently associated with adverse effects, including gastrointestinal disturbances, insomnia, weight gain, dizziness, and sexual dysfunction, which may reduce patient compliance and treatment success.12
These limitations have led to growing interest in alternative and complementary therapeutic approaches. In this context, medicinal plants have attracted considerable attention as potential sources of novel antidepressant agents.17 Traditional systems of medicine have long utilized plant-based remedies for the treatment of neurological and psychological disorders.25 Medicinal plants are rich in bioactive phytochemicals such as flavonoids, alkaloids, terpenoids, glycosides, and phenolic compounds, many of which exhibit antioxidant, anti-inflammatory, and neuroprotective properties.26,31 These phytoconstituents may exert antidepressant-like effects through multiple mechanisms, including modulation of monoaminergic neurotransmission, reduction of oxidative stress, suppression of neuroinflammatory responses, and enhancement of neurogenesis and synaptic plasticity.25,30 (Fig. 1 illustrates the major neurobiological factors involved in depression.).
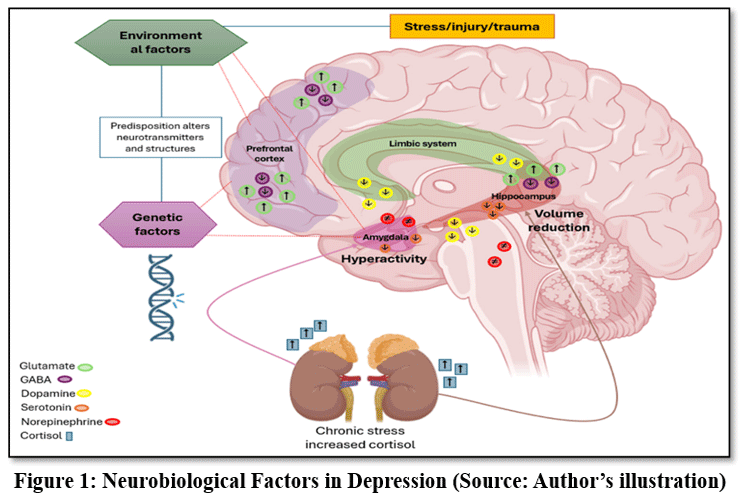 |
Figure 1: Neurobiological Factors in Depression (Source: Author’s illustration).
|
Experimental animal models play a crucial role in understanding the neurobiological basis of depression and in evaluating the antidepressant potential of plant-derived compounds.14 These models provide controlled environments for studying behavioral, biochemical, and neurochemical alterations associated with depressive states. Widely used models include the forced swim test, tail suspension test, chronic unpredictable mild stress model, and reserpine-induced depression model.14,16 Each of these models reflects different aspects of depressive pathology and allows researchers to assess the pharmacological effects of therapeutic agents.36 Consequently, preclinical studies using these models represent an essential step in the development of plant-based antidepressant therapies.
Pathophysiology of Depression
Depression is a multifactorial neuropsychiatric disorder that arises from complex interactions among genetic susceptibility, environmental stressors, and neurobiological alterations.13 The underlying mechanisms involve disturbances in neurotransmission, inflammatory signaling, oxidative balance, endocrine regulation, and neuronal plasticity. Rather than being attributed to a single cause, depression is now considered the result of dysregulation across multiple interconnected biological systems that influence brain structure and function.
Advances in neuroscience and molecular biology have provided deeper insight into the neurobiological processes involved in depressive disorders. Several hypotheses have been proposed to explain the pathogenesis of depression, including alterations in monoaminergic neurotransmission, neuroinflammatory responses, oxidative stress, dysregulation of the hypothalamic–pituitary–adrenal (HPA) axis, and impairment of neuroplasticity.14 These mechanisms collectively contribute to structural and functional changes in brain regions involved in mood regulation, cognition, and emotional processing, such as the hippocampus, prefrontal cortex, and amygdala. (The mechanisms described are summarized in Fig. 2.)
Monoamine Hypothesis
The monoamine hypothesis is one of the earliest and most extensively studied theories explaining the biological basis of depression.15 According to this hypothesis, depressive symptoms arise from deficiencies or dysregulation of monoamine neurotransmitters in the central nervous system, particularly serotonin (5-hydroxytryptamine), norepinephrine, and dopamine.16 These neurotransmitters play essential roles in regulating mood, motivation, reward processing, and emotional stability.
Serotonin is primarily involved in the regulation of mood, sleep, appetite, and emotional behavior. Reduced serotonergic neurotransmission has been associated with increased susceptibility to depressive symptoms, including anxiety, irritability, and emotional instability. Similarly, norepinephrine contributes to alertness, attention, and the body’s response to stress. A decrease in norepinephrine signaling may lead to fatigue, reduced concentration, and diminished motivation. Dopamine, another critical monoamine neurotransmitter, is closely linked to reward pathways and pleasure perception, and its impairment is associated with anhedonia, a core feature of depression.17
Alterations in monoamine levels may occur through various mechanisms, including reduced synthesis, increased enzymatic degradation, or enhanced reuptake by presynaptic neurons. These changes disrupt synaptic communication and impair normal neural signaling pathways. The monoamine hypothesis is supported by the pharmacological action of antidepressant medications such as selective serotonin reuptake inhibitors (SSRIs) and monoamine oxidase inhibitors (MAOIs), which increase monoamine availability in the synaptic cleft.18 However, this hypothesis alone does not fully explain the complexity of depression, indicating the involvement of additional mechanisms.
Neuroinflammation
Increasing evidence suggests that neuroinflammatory processes play a crucial role in the development and progression of depression.19 Neuroinflammation involves activation of immune responses within the central nervous system, primarily mediated by microglia and astrocytes. These glial cells regulate immune activity in the brain and release inflammatory mediators in response to stress or injury.
In depressive conditions, elevated levels of pro-inflammatory cytokines such as tumor necrosis factor-alpha (TNF-α), interleukin-1 beta (IL-1β), and interleukin-6 (IL-6) have been observed. These cytokines can interfere with neurotransmitter metabolism, synaptic plasticity, and neuronal survival. For example, inflammatory mediators may alter tryptophan metabolism, reducing serotonin synthesis. Chronic neuroinflammation can also disrupt neuronal communication and contribute to structural damage in brain regions associated with mood regulation.20
Microglial activation is a key feature of neuroinflammation in depression. While microglia normally support neuronal homeostasis and synaptic remodeling, persistent activation leads to excessive production of inflammatory molecules and reactive oxygen species, which impair neuronal function and promote neurodegeneration.
Oxidative Stress
Oxidative stress is an important factor in the pathophysiology of depression and occurs when there is an imbalance between the production of reactive oxygen species (ROS) and the antioxidant defense system.21 Excessive ROS can damage lipids, proteins, and nucleic acids, leading to cellular dysfunction.
The brain is particularly vulnerable to oxidative damage due to its high oxygen consumption and lipid-rich composition. Increased levels of oxidative stress markers have been reported in individuals with depression, indicating enhanced neuronal damage. Oxidative stress can impair neurotransmission, disrupt membrane integrity, and alter mitochondrial function, ultimately affecting neuronal survival and function.
Furthermore, oxidative stress interacts with other pathological mechanisms such as inflammation and neurotransmitter imbalance, thereby contributing to the overall progression of depressive disorders.
Hypothalamic–Pituitary–Adrenal (HPA) Axis Dysfunction
The hypothalamic–pituitary–adrenal axis is a central neuroendocrine system that regulates the body’s response to stress.22 Under normal conditions, activation of this system leads to the release of corticotropin-releasing hormone from the hypothalamus, which stimulates adrenocorticotropic hormone release from the pituitary gland, ultimately leading to cortisol secretion from the adrenal cortex.
In depression, dysregulation of the HPA axis is frequently observed, particularly in response to chronic stress. Persistent activation of this system results in prolonged elevation of cortisol levels, which can have detrimental effects on brain function. Elevated glucocorticoids are associated with reduced neurogenesis, neuronal atrophy, and impaired synaptic connectivity in key brain regions such as the hippocampus.
Additionally, disruption of feedback mechanisms that normally regulate HPA axis activity leads to sustained hormonal imbalance, further contributing to depressive symptoms and increasing the risk of recurrence.
Neuroplasticity Impairment
Neuroplasticity refers to the brain’s ability to adapt and reorganize in response to environmental stimuli, learning, and experience.23 It involves processes such as synaptic remodeling, neuronal growth, and neurogenesis.
In depression, neuroplasticity is significantly impaired, particularly in brain regions involved in mood regulation, such as the hippocampus and prefrontal cortex. Reduced synaptic connectivity and decreased neuronal survival contribute to cognitive deficits and emotional dysregulation.
Brain-derived neurotrophic factor (BDNF) is a key molecule involved in neuroplasticity, supporting neuronal survival, growth, and synaptic formation. Decreased levels of BDNF have been observed in individuals with depression and in experimental models. Restoration of BDNF levels has been associated with improved neuronal function and is considered an important mechanism underlying the therapeutic effects of antidepressants.24
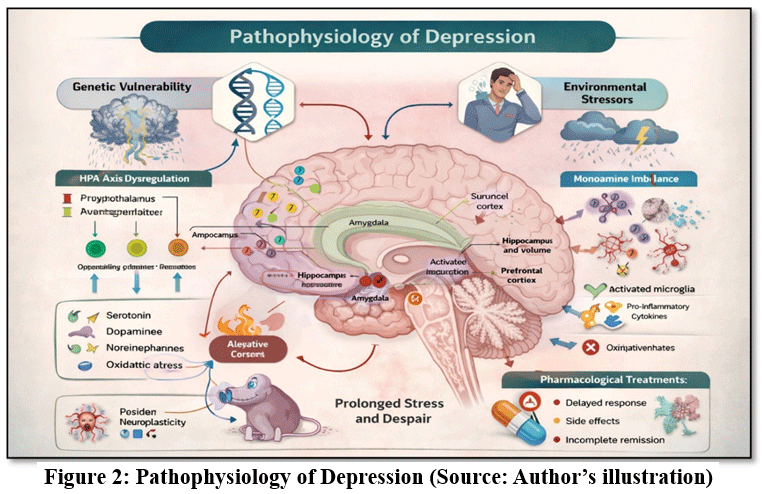 |
Figure 2: Pathophysiology of Depression (Source: Author’s illustration)
|
Experimental Animal Models of Depression
Understanding the biological mechanisms underlying depression and evaluating the therapeutic potential of novel antidepressant agents requires reliable experimental models.23 Animal models play a crucial role in preclinical research by enabling controlled investigation of behavioral, biochemical, and neurophysiological changes associated with depressive states. Although human depression is a complex and heterogeneous condition, several behavioral paradigms in rodents have been developed to replicate specific symptoms or biological aspects of the disorder.24 These models provide valuable platforms for studying disease mechanisms and for screening pharmacological as well as plant-derived compounds with potential antidepressant activity.
Experimental models of depression are generally designed to reproduce behavioral features such as behavioral despair, anhedonia, reduced motivation, and stress-induced behavioral alterations. In addition to behavioral outcomes, these models also allow the investigation of neurochemical changes, endocrine responses, and structural alterations in the brain.25 Among the most widely used models in preclinical depression research are the forced swim test, tail suspension test, chronic unpredictable mild stress model, and reserpine-induced depression model. Each of these models reflects different aspects of depressive pathology and offers specific advantages for evaluating antidepressant-like effects. (Common experimental models are presented in Fig. 3.)
Forced Swim Test (FST)
The forced swim test is one of the most commonly used behavioral models for assessing antidepressant activity in rodents.26 This model is based on the concept of behavioral despair and evaluates the tendency of an animal to cease active escape-directed behaviors when exposed to an inescapable stress condition.
In this test, rodents are placed individually in a transparent cylindrical container filled with water maintained at a controlled temperature. Initially, the animals display active behaviors such as swimming and climbing in an attempt to escape. Over time, these efforts decrease, and the animals adopt an immobile posture, making only minimal movements to keep their heads above water.
The duration of immobility is considered an indicator of depressive-like behavior and is widely used as a quantitative measure in antidepressant screening. Compounds with antidepressant properties typically reduce immobility time and increase active behaviors. Interestingly, different classes of antidepressants may influence specific behavioral patterns, with serotonergic agents increasing swimming behavior and noradrenergic agents enhancing climbing activity.
In addition to behavioral assessment, the forced swim test has also been used to study neurochemical and hormonal changes associated with stress exposure. Due to its simplicity, reproducibility, and sensitivity, it remains a widely accepted method for evaluating antidepressant potential.
Tail Suspension Test (TST)
The tail suspension test is another widely used behavioral model for screening antidepressant activity, particularly in mice.27 Similar to the forced swim test, this model is based on the principle of behavioral despair and evaluates the inability of the animal to cope with an unavoidable stress condition.
In this procedure, mice are suspended by their tails using adhesive tape attached to a horizontal support. Initially, the animals exhibit vigorous movements such as swinging and twisting as they attempt to escape. After repeated unsuccessful attempts, they become immobile, displaying minimal movements necessary for balance.
The duration of immobility is recorded as an indicator of depressive-like behavior. Antidepressant agents typically reduce immobility time and increase active escape-oriented movements. This test is relatively simple to perform and does not require complex equipment, making it suitable for rapid screening of multiple compounds.
The tail suspension test is highly sensitive to a variety of antidepressant drugs, particularly those affecting monoaminergic systems. It also serves as a complementary model to the forced swim test, providing additional insights into behavioral responses under stress.
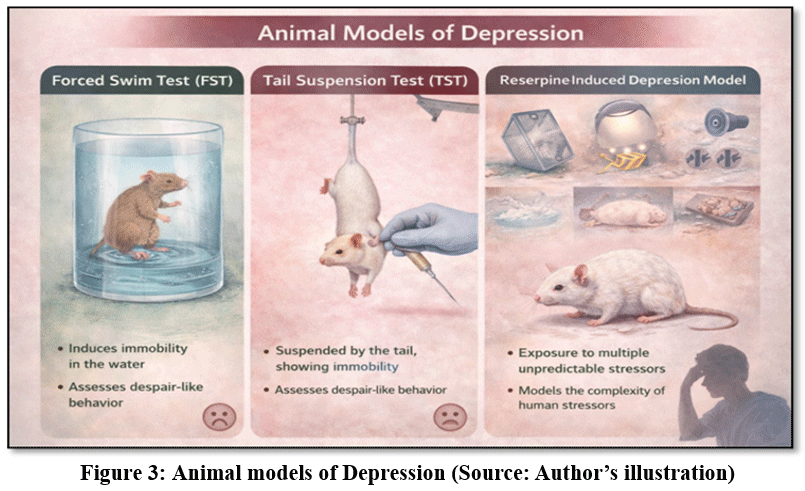 |
Figure 3: Animal models of Depression (Source: Author’s illustration)
|
Chronic Unpredictable Mild Stress Model (CUMS)
The chronic unpredictable mild stress model is considered one of the most relevant experimental paradigms for studying depression, as it closely mimics the effects of prolonged exposure to stress.28 Unlike acute behavioral models, this approach focuses on long-term behavioral and physiological changes induced by chronic stress.
In this model, animals are exposed to a series of mild stressors applied in an unpredictable sequence over an extended period. These stressors may include alterations in light–dark cycles, food or water deprivation, cage tilting, wet bedding, and social isolation. The unpredictability prevents adaptation, resulting in sustained stress responses.
One of the key behavioral outcomes observed in this model is anhedonia, which is commonly assessed using the sucrose preference test. Animals subjected to chronic stress exhibit reduced preference for sweet solutions, reflecting decreased reward sensitivity. Additional changes may include reduced locomotor activity, altered sleep patterns, and decreased exploratory behavior.
At the neurobiological level, chronic stress induces significant alterations in neurotransmitter systems, inflammatory pathways, endocrine responses, and brain structure. Due to its ability to replicate multiple features of depression, the CUMS model is widely used for evaluating long-term antidepressant effects.
Reserpine-Induced Depression Model
The reserpine-induced model is a pharmacological approach used to study depressive-like behavior in experimental animals.29 Reserpine is an alkaloid that interferes with the storage of monoamine neurotransmitters by inhibiting vesicular monoamine transporters, leading to depletion of serotonin, dopamine, and norepinephrine in the brain.
Administration of reserpine produces behavioral and physiological changes that resemble depressive symptoms, including reduced locomotor activity, ptosis, hypothermia, and increased immobility. These effects reflect disruption of monoaminergic signaling pathways involved in mood regulation.
This model is particularly useful for studying the role of neurotransmitter depletion in depression and for evaluating compounds that restore monoamine levels. Many antidepressant drugs and plant-derived extracts have been shown to reverse reserpine-induced behavioral changes, indicating their potential therapeutic effects.30 Additionally, biochemical analyses in this model often include measurement of neurotransmitter levels and enzymatic activity, providing insight into the mechanisms underlying antidepressant action.
Table 1: Animal Model Comparison in Depression
| Sr. no | Animal Models | Features | Advantages |
| 1 | Forced Swim Test (FST) | Measures behavioral despair (immobility time).18,26 | Simple, rapid, widely used screening model |
| 2 | Tail Suspension Test (TST) | Assesses stress-induced immobility.17,27 | High sensitivity to antidepressants |
| 3 | Chronic Unpredictable Mild Stress (CUMS) | Mimics chronic stress and anhedonia.14,28 | Closely resembles human depression |
| 4 | Reserpine-Induced Model | Depletes monoamine neurotransmitters.19,29 | Useful for studying the monoamine hypothesis |
Medicinal Plants with Antidepressant Activity
Medicinal plants exert antidepressant effects through multi-target mechanisms that collectively regulate neurochemical and cellular processes involved in depression. Unlike conventional drugs that often act on single pathways, phytochemicals influence interconnected biological systems.3
Table 2: Medicinal plants reported to exhibit antidepressant activity in experimental animal models
| No. | Medicinal Plant | Family | Experimental Model | Key Active Compounds | Proposed Mechanism of Antidepressant Action |
| 1 | Hypericum perforatum | Hypericaceae | FST, TST | Hypericin, Hyperforin | Inhibits monoamine reuptake and enhances synaptic availability of serotonin and dopamine.29 |
| 2 | Withania somnifera | Solanaceae | CUMS, FST | Withanolides | Reduces stress-induced endocrine disturbances and promotes neuronal resilience.26 |
| 3 | Bacopa monnieri | Plantaginaceae | FST, TST | Bacosides | Enhances synaptic signaling and improves neuronal communication involved in mood regulation.27 |
| 4 | Curcuma longa | Zingiberaceae | CUMS, TST | Curcumin | Suppresses inflammatory signaling pathways and protects neuronal structures.25 |
| 5 | Panax ginseng | Araliaceae | FST | Ginsenosides | Enhances neural energy metabolism and modulates central neurotransmitter systems.21 |
| 6 | Ginkgo biloba | Ginkgoaceae | CUMS, FST | Flavone glycosides | Improves cerebral circulation and stabilizes neuronal oxidative balance.21 |
Hypericum perforatum (St. John’s Wort)
Plant Extract
Hypericum perforatum is a well-known medicinal herb traditionally used for treating mood disorders. Hydroalcoholic and ethanolic extracts of the aerial parts of the plant are commonly used in experimental studies. These extracts contain several pharmacologically active constituents, including hypericin, hyperforin, flavonoids, and phenolic compounds. Hyperforin in particular has been identified as a key compound responsible for the antidepressant properties of the plant due to its ability to influence neuronal signaling pathways.
Animal Model Used
The antidepressant activity of Hypericum perforatum has been extensively evaluated using behavioral models such as the forced swim test (FST) and tail suspension test (TST) in rodents. These models assess behavioral despair by measuring immobility time when animals are exposed to an inescapable stressful situation.
Dose: In most experimental investigations, the plant extract is administered orally or intraperitoneally at doses typically ranging between 100 and 300 mg/kg body weight in rodents.
Observed Antidepressant Effect
Administration of Hypericum perforatum extract has been shown to significantly reduce immobility time in both the forced swim and tail suspension tests, indicating an improvement in stress-coping behavior. The treatment enhances locomotor activity and increases escape-oriented behaviors in experimental animals. Neurochemical studies suggest that these behavioral improvements are associated with increased synaptic availability of serotonin, dopamine, and norepinephrine in the brain. Additionally, the extract may modulate intracellular signaling pathways linked to mood regulation and improve neuronal resilience against stress-induced functional disturbances.32
Withania somnifera (Ashwagandha)
Plant Extract
Withania somnifera, commonly known as Ashwagandha, is widely used in traditional medicine for its adaptogenic and neuroprotective properties. Experimental studies typically utilize aqueous or ethanolic extracts of the roots, which are rich in steroidal lactones known as withanolides. These compounds contribute to the plant’s ability to influence physiological responses to stress.
Animal Model Used
The antidepressant potential of Withania somnifera has been studied primarily using chronic unpredictable mild stress (CUMS) models and the forced swim test in rodents. The CUMS model is particularly valuable because it reproduces behavioral changes associated with prolonged stress exposure.
Dose
Preclinical studies have generally used doses ranging from 50 to 200 mg/kg body weight administered orally over several days or weeks.
Observed Antidepressant Effect
Treatment with Withania somnifera extract has demonstrated the ability to improve behavioral performance in stressed animals. Rodents receiving the extract exhibit increased mobility and enhanced exploratory activity compared with untreated stressed animals. In chronic stress models, the extract restores normal preference for sucrose solutions, indicating improvement in anhedonia-like symptoms. These behavioral effects are accompanied by normalization of stress-related hormonal responses and improved neuronal adaptability within brain regions involved in emotional processing. The plant’s adaptogenic nature appears to support physiological resilience during prolonged stress exposure.33
Curcuma longa (Turmeric)
Plant Extract
Curcuma longa, commonly referred to as turmeric, contains the polyphenolic compound curcumin, which is responsible for many of its pharmacological properties. Ethanolic extracts of turmeric rhizomes are frequently used in experimental research due to their high concentration of curcuminoids.
Animal Model Used
Antidepressant-like activity of turmeric extracts has been investigated using tail suspension tests, forced swim tests, and chronic stress paradigms in rodents.
Dose
Curcumin or turmeric extract is typically administered at doses between 50 and 200 mg/kg body weight in experimental animals.
Observed Antidepressant Effect
Experimental findings indicate that treatment with Curcuma longa extract reduces passive immobility behavior in rodents exposed to stressful conditions. Animals receiving curcumin demonstrate improved activity patterns and enhanced motivation to escape stressful environments. In addition to behavioral improvements, biochemical analyses have shown that turmeric extract can influence neural signaling pathways associated with mood regulation. Curcumin also appears to enhance neuronal survival by maintaining cellular defense mechanisms within the brain. These properties suggest that turmeric may exert antidepressant effects by supporting neuronal stability and improving neural communication networks involved in emotional regulation.34
Bacopa monnieri
Plant Extract
Bacopa monnieri, also known as Brahmi, is a traditional medicinal plant recognized for its cognitive-enhancing and neuroprotective effects. Extracts used in research are typically prepared from the whole plant, containing bioactive saponins collectively known as bacosides.
Animal Model Used
The antidepressant-like activity of Bacopa monnieri has been evaluated using forced swim tests, tail suspension tests, and chronic stress-induced behavioral models.
Dose
Experimental studies commonly administer plant extracts in the range of 40 to 120 mg/kg body weight in rodents.
Observed Antidepressant Effect
Administration of Bacopa monnieri extract results in a noticeable reduction in immobility time during behavioral tests, suggesting improved coping behavior in stressful situations. Animals treated with the extract show increased locomotor activity and enhanced behavioral responsiveness. Additionally, the plant has been associated with improvements in cognitive performance and memory functions in stressed animals. These effects may be attributed to the ability of bacosides to support neuronal communication and maintain the structural integrity of neural networks. By strengthening synaptic signaling processes, Bacopa monnieri contributes to improved emotional and cognitive functioning.35
Vitex negundo
Plant Extract
Vitex negundo is a medicinal shrub widely used in traditional medicine for treating inflammatory and neurological conditions. Experimental research generally employs methanolic or ethanolic extracts of the leaves, which contain flavonoids, iridoid glycosides, and other phenolic compounds.
Animal Model Used
The antidepressant potential of Vitex negundo has been investigated using reserpine-induced depression models, as well as forced swim tests in rodents.
Dose
Studies typically administer the extract at doses between 100 and 400 mg/kg body weight, depending on the experimental design.
Observed Antidepressant Effect
Rodents treated with Vitex negundo extract exhibit marked improvement in behavioral performance compared with untreated animals exposed to depressive stimuli. The extract significantly reduces immobility behavior and restores normal locomotor patterns in reserpine-treated animals. Biochemical investigations suggest that the plant extract may help restore normal neurotransmitter activity that is disrupted by pharmacological depletion. Furthermore, its phytochemical constituents appear to protect neuronal tissues from stress-related biochemical disturbances, thereby supporting normal brain function and behavioral stability.36
Other Medicinal Plants
Several other medicinal plants, including Panax ginseng, Ginkgo biloba, Rhodiola rosea, Centella asiatica, Ocimum sanctum, and Valeriana officinalis, have also demonstrated antidepressant-like effects in experimental models.37 These plants exert their effects through various mechanisms such as modulation of neurotransmitter systems, enhancement of antioxidant defenses, and improvement of neuronal resilience under stress conditions.
Overall, medicinal plants represent a diverse and promising source of bioactive compounds with potential antidepressant properties. Their ability to act on multiple biological targets makes them valuable candidates for the development of alternative therapeutic strategies in the management of depression.38
Mechanisms of Antidepressant Action of Medicinal Plants
Medicinal plants contain a wide range of bioactive phytochemicals that act on multiple biological pathways involved in mood regulation. Unlike conventional antidepressants, which generally target a single neurotransmitter system, plant-derived compounds often exhibit multi-targeted actions that influence neurochemical balance, oxidative stress, inflammatory responses, and neuroendocrine regulation.39
Table 3: Mechanisms vs Phytochemicals
| Sr.No | Mechanism of Action | Phytochemical compounds | Example Medicinal plants |
| 1 | Modulation of monoamine neurotransmitters | Hypericin, Hyperforin, Bacosides.27,29 | Curcuma longa, Ginkgo biloba |
| 2 | Anti-inflammatory effects | Curcumin, Withanolides.25,26 | Curcuma longa, Withania somnifera |
| 3 | Antioxidant activity | Flavonoids, Polyphenols, Curcumin.25,31 | Curcuma longa, Ginkgo biloba |
| 4 | Neuroprotection | Asiaticoside, Ginsenosides.23,24 | Centella asiatica, Panax ginseng |
| 5 | Regulation of HPA axis | Withanolides, Rosavin.22,26 | Withania somnifera, Rhodiola rosea |
| 6 | Enhancement of neuroplasticity (BDNF) | Bacosides, Flavonoids.27,34 | Bacopa monnieri, Ginkgo biloba |
Modulation of Neurotransmitter Systems
One of the primary mechanisms through which medicinal plants exert antidepressant effects is the modulation of monoaminergic neurotransmitter systems.
Several plant-derived compounds regulate the synthesis, release, and availability of key neurotransmitters such as serotonin, dopamine, and norepinephrine. These effects are achieved through inhibition of monoamine oxidase enzymes, modulation of transporter proteins, and enhancement of presynaptic neurotransmitter release. By restoring synaptic neurotransmitter balance, phytochemicals improve neural communication within brain regions responsible for mood and emotional processing.40
Antioxidant Activity
Oxidative damage plays a critical role in neuronal dysfunction associated with depression. Phytochemicals such as flavonoids and phenolic compounds act as potent free radical scavengers, reducing lipid peroxidation and protecting cellular macromolecules. In addition, these compounds enhance endogenous antioxidant systems, including superoxide dismutase and glutathione pathways, thereby maintaining redox homeostasis in neuronal tissues.41
Anti-Inflammatory Effects
Chronic neuroinflammation contributes to the progression of depressive disorders by disrupting neurotransmitter metabolism and neuronal signaling. Medicinal plants inhibit the activation of microglial cells and reduce the release of pro-inflammatory cytokines such as IL-6 and TNF-α. This anti-inflammatory action helps restore a balanced neuroimmune environment and prevents further neuronal damage.42
Neuroprotective Effects
Impaired neuroplasticity is a hallmark of depression. Phytochemicals promote neuronal growth, synaptic remodeling, and neurogenesis by upregulating neurotrophic factors such as brain-derived neurotrophic factor (BDNF). These effects improve synaptic connectivity and strengthen neural circuits involved in cognitive and emotional regulation.43
Regulation of the HPA Axis
Prolonged activation of the hypothalamic–pituitary–adrenal axis results in elevated cortisol levels, which negatively affect neuronal integrity. Adaptogenic medicinal plants help normalize HPA axis activity by reducing stress hormone levels and improving feedback regulation. This contributes to improved stress resilience and stabilization of mood-related neuroendocrine functions.44,45
Histopathological Evidence Supporting the Antidepressant Potential of Medicinal Plants
Histopathological investigations provide essential insights into the structural and cellular alterations that occur in the brain during depressive disorders. Chronic stress and chemically induced models of depression are known to produce significant morphological changes in neuronal tissues, particularly in brain regions involved in emotional regulation and cognitive processing.39 Examination of brain sections obtained from experimental animals has consistently shown that treatment with various medicinal plant extracts can significantly improve neuronal integrity and reduce pathological alterations associated with depressive conditions.40 (Representative histopathological changes are shown in Fig. 4.) These histological findings strongly support the concept that plant-derived phytochemicals possess considerable neuroprotective potential.
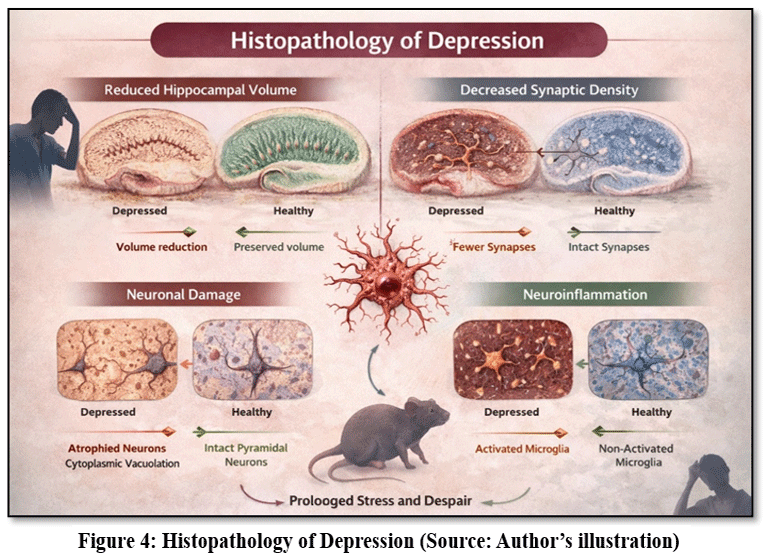 |
Figure 4: Histopathology of Depression (Source: Author’s illustration)
|
Preservation of Hippocampal Pyramidal Neurons
The hippocampus is among the most sensitive brain regions affected by chronic stress and depressive pathology. Within this structure, pyramidal neurons play a crucial role in processes such as learning, memory formation, and emotional regulation.41 Histological observations in animal models of depression frequently demonstrate shrinkage, disorganization, and degeneration of these neurons, particularly in the CA1 and CA3 regions of the hippocampus. These structural abnormalities are often accompanied by reduced synaptic connectivity and impaired neuronal signaling.
Experimental studies involving treatment with medicinal plant extracts have demonstrated significant improvement in hippocampal architecture. Microscopic examination of brain tissues from treated animals reveals well-preserved pyramidal neuronal layers, enhanced cellular organization, and more distinct nuclear morphology when compared to untreated depressed animals.42 These observations indicate that phytochemicals present in medicinal plants may protect hippocampal neurons from stress-induced structural damage while also supporting the maintenance of normal neuronal function.
Reduction of Microglial Activation
Microglial cells function as the primary immune defense within the central nervous system and become activated in response to neuronal injury or inflammatory stimuli. In experimental models of depression, excessive activation of microglia is commonly observed, leading to the release of inflammatory mediators that further contribute to neuronal damage and disruption of neural communication.43
Histopathological evaluations have shown that animals exposed to chronic stress or pharmacologically induced depression exhibit an increased number of activated microglial cells across various brain regions. These activated cells typically appear enlarged and display morphological characteristics associated with inflammatory responses. Treatment with specific medicinal plant extracts has been found to significantly reduce this microglial activation. Brain sections from treated animals demonstrate fewer activated microglial cells along with restoration of more normal cellular morphology, suggesting that plant-derived compounds may play an important role in re-establishing a balanced neuroimmune environment within the brain.
Enhancement of Neuronal Density
Depression is frequently associated with a reduction in neuronal density in key brain regions involved in mood regulation. Prolonged exposure to stress can lead to neuronal loss or decreased neuronal proliferation, resulting in functional impairment of neural circuits.44 Histological staining methods often reveal a reduced number of intact neurons in areas such as the hippocampus and cerebral cortex in untreated depressed animals.
Experimental findings indicate that administration of medicinal plant extracts can significantly enhance neuronal density in these regions. Animals treated with phytochemical-rich extracts typically exhibit a higher number of healthy neuronal cells along with improved cellular distribution when compared to untreated groups. This increase in neuronal population may be attributed to the ability of plant-derived compounds to support neuronal survival and create favorable conditions for maintaining cellular integrity within neural tissues.
Prevention of Degenerative Changes in the Cerebral Cortex
The cerebral cortex is another critical brain region responsible for higher cognitive functions, decision-making processes, and emotional regulation. In depressive conditions, histopathological examination often reveals degenerative alterations such as neuronal shrinkage, cytoplasmic vacuolation, and disruption of cortical layering.45 These structural changes can interfere with communication between cortical and limbic regions of the brain, thereby contributing to behavioral and emotional disturbances observed in depression.
Treatment with medicinal plant extracts has been reported to effectively reduce these pathological changes. Histological sections of cortical tissue from treated animals commonly show better preservation of cortical layers, improved neuronal morphology, and reduced signs of degeneration. These structural improvements suggest that phytochemicals may help stabilize neuronal membranes and protect cortical neurons from stress-induced damage, thereby supporting overall brain function.
Future Perspectives in the Development of Plant-Based Antidepressant Therapies
Although extensive preclinical investigations have demonstrated the antidepressant potential of numerous medicinal plants, several scientific and methodological challenges must be addressed before these findings can be effectively translated into clinical practice. Advancing research in this field requires a multidisciplinary approach that integrates pharmacology, phytochemistry, neuroscience, and clinical medicine. Addressing the current limitations will not only improve the reliability of experimental outcomes but also facilitate the development of safe and effective plant-derived therapeutic agents for depressive disorders.
Standardization of Medicinal Plant Extracts
One of the primary challenges in phytopharmacological research is the lack of standardized plant preparations. The chemical composition of herbal extracts can vary widely depending on factors such as geographical origin, climate conditions, harvesting time, and extraction techniques. As a result, different studies investigating the same plant species may report inconsistent pharmacological effects. This variability complicates the interpretation of experimental findings and makes it difficult to establish reproducible therapeutic outcomes.
Future investigations should prioritize the development of standardized extraction protocols and quality control measures. Advanced analytical techniques, including chromatographic and spectroscopic methods, can be used to identify and quantify key bioactive constituents within plant extracts. Establishing consistent phytochemical profiles will help ensure that experimental treatments contain defined concentrations of active compounds. Such standardization is essential for improving experimental reproducibility and for facilitating regulatory approval of plant-based medicinal products.
Expansion of Clinical Research
While animal models have provided valuable insights into the antidepressant potential of medicinal plants, the translation of these findings into human clinical settings remains limited. Only a small number of herbal preparations have undergone rigorous clinical evaluation for the treatment of depressive disorders. Without well-designed clinical trials, it is difficult to determine the safety, efficacy, and optimal dosage of plant-derived therapies in human populations.
Future research efforts should therefore focus on conducting controlled clinical studies involving diverse patient populations. These investigations should evaluate not only the therapeutic effectiveness of plant extracts but also their long-term safety profiles and potential interactions with conventional antidepressant medications. Integrating clinical research with pharmacokinetic and pharmacodynamic studies will provide a clearer understanding of how plant-derived compounds behave within the human body. Such information is crucial for establishing evidence-based guidelines for the clinical use of herbal antidepressants.
Understanding Phytochemical Variability
Another important consideration in medicinal plant research is the complex and dynamic nature of phytochemical composition. Individual plant species often contain hundreds of chemical constituents, many of which may contribute synergistically to their therapeutic effects. Environmental conditions, soil composition, plant maturity, and processing methods can influence the concentration and stability of these compounds.
Future studies should employ advanced metabolomic and molecular profiling techniques to better understand the diversity of phytochemicals present in medicinal plants. By identifying the specific compounds responsible for antidepressant activity, researchers can gain deeper insights into their mechanisms of action. Such knowledge may also support the development of optimized plant formulations or the isolation of novel therapeutic molecules. Investigating potential synergistic interactions among phytochemicals may further enhance the effectiveness of plant-based treatments.
Integration of Translational Research Approaches
Bridging the gap between experimental research and clinical application represents an important objective for future studies in this field. Translational research strategies that combine behavioral neuroscience, molecular biology, and clinical pharmacology will be essential for advancing plant-derived antidepressant therapies. Investigations exploring gene expression changes, neural signaling pathways, and brain imaging techniques may provide valuable insights into how phytochemicals influence neural networks associated with mood regulation.
Additionally, emerging technologies such as nanotechnology-based drug delivery systems may improve the bioavailability and therapeutic efficiency of plant-derived compounds. Incorporating such innovative approaches could significantly enhance the clinical potential of medicinal plants and allow more precise targeting of neurobiological pathways involved in depression.
Discussion
Depression is a multifactorial neuropsychiatric disorder characterized by complex interactions among neurochemical, inflammatory, endocrine, and structural alterations in the brain. Although conventional antidepressants primarily target monoaminergic systems, their delayed onset of action and limited efficacy in some patients necessitate the exploration of alternative therapeutic approaches. In this context, medicinal plants have emerged as promising candidates due to their diverse phytochemical composition and ability to modulate multiple biological pathways simultaneously.
Preclinical studies provide substantial evidence that plant-derived compounds exert antidepressant-like effects through integrated mechanisms. These include modulation of key neurotransmitter systems, enhancement of neuronal resilience, and regulation of stress-related signaling pathways in brain regions such as the hippocampus, prefrontal cortex, and amygdala. Unlike single-target conventional drugs, phytochemicals often act on interconnected pathways, contributing to improved neural communication and adaptive responses to stress.
In addition to neurochemical modulation, medicinal plants play a crucial role in maintaining cellular homeostasis within the brain. Their antioxidant properties help counteract oxidative damage, while anti-inflammatory effects reduce neuroimmune activation that can impair neuronal function. These combined actions support neuronal integrity and preserve synaptic function under chronic stress conditions.
Structural and functional improvements in the brain further support the therapeutic potential of plant-based interventions. Experimental findings indicate that medicinal plant extracts can protect against neuronal damage, maintain cellular organization, and promote neuroplasticity. Enhancement of neurotrophic factors, particularly those involved in synaptic remodeling and neuronal survival, appears to be a key mechanism underlying long-term antidepressant effects.
Animal models of depression continue to serve as valuable tools for evaluating these effects, enabling the assessment of behavioral, biochemical, and structural outcomes. Consistent findings across different experimental paradigms strengthen the evidence for the antidepressant potential of medicinal plants.
Despite these promising observations, certain limitations must be considered. Variability in phytochemical composition, differences in extraction methods, and reliance on preclinical models may affect the reproducibility and clinical translation of findings. Therefore, standardization of plant extracts and well-designed clinical studies are essential to establish their safety, efficacy, and therapeutic applicability in humans.
Overall, medicinal plants represent a rich source of bioactive compounds with multi-targeted actions. Their ability to address the complex and interconnected mechanisms underlying depression highlights their potential as complementary or alternative therapeutic options in the management of depressive disorders.
Future research should focus on well-designed clinical trials to validate the efficacy and safety of medicinal plant-derived compounds in human populations. Integration of pharmacokinetic, pharmacodynamic, and molecular studies will be essential to facilitate the translation of preclinical findings into clinically applicable antidepressant therapies.
Limitations
Despite significant progress in preclinical research, several limitations must be considered when interpreting the antidepressant potential of medicinal plants. Animal models of depression, although widely used, do not fully replicate the complexity and heterogeneity of human depressive disorders. Behavioral tests such as the forced swim test and tail suspension test primarily assess acute stress responses and may not accurately reflect chronic emotional and cognitive symptoms observed in humans. Additionally, variations in experimental design, species differences, and phytochemical composition of plant extracts can influence study outcomes. Furthermore, most evidence is derived from preclinical studies, and there is a lack of well-controlled clinical trials to validate these findings in human populations. These limitations highlight the need for standardized methodologies and translational research approaches to bridge the gap between experimental studies and clinical application.
Conclusion
Depression remains a major global health challenge due to its complex pathophysiology and the limitations associated with currently available pharmacological treatments. The growing body of experimental evidence highlights the significant therapeutic potential of medicinal plants in the management of depressive disorders. Plant-derived phytochemicals possess diverse pharmacological properties that can influence several biological pathways involved in mood regulation, including neurotransmitter modulation, antioxidant defense, anti-inflammatory activity, neuroprotection, and neuroendocrine regulation.
Preclinical studies employing validated animal models of depression have consistently demonstrated the ability of various medicinal plant extracts to alleviate depressive-like behaviors and improve neurobiological parameters. These effects are often accompanied by preservation of neuronal structure, reduction of neuroinflammatory responses, and enhancement of neuroplastic processes within critical brain regions such as the hippocampus and cerebral cortex. The multifactorial actions of phytochemicals provide a promising approach for targeting the complex biological mechanisms underlying depressive disorders.
Nevertheless, significant challenges remain before plant-based antidepressant therapies can be widely implemented in clinical practice. Variability in phytochemical composition, lack of standardized extraction methods, and limited clinical validation represent major obstacles in the development of reliable herbal treatments. Future research should focus on the identification of active phytoconstituents, elucidation of molecular mechanisms, and well-designed clinical trials to confirm the safety and efficacy of medicinal plants in the treatment of depression.
However, translation of preclinical findings into clinical practice requires rigorous human trials and standardization of herbal formulations. In conclusion, medicinal plants represent a promising reservoir of novel neuroactive compounds with the potential to contribute to the development of safer and more effective antidepressant therapies. Continued interdisciplinary research integrating pharmacology, neuroscience, and phytochemistry will be essential for translating preclinical findings into clinically applicable treatments for depressive disorders.
The findings of this review suggest that medicinal plants and their bioactive phytochemicals hold significant potential as multi-target therapeutic agents, offering a promising complementary or alternative approach to conventional antidepressant treatments in clinical practice.
Acknowledgement
The authors would like to acknowledge the Department of Pharmacology, Navsahyadri Institute of Pharmacy, for providing the necessary facilities to conduct this review.
Funding Sources
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Conflict of Interest
The authors do not have any conflict of interest.
Data Availability Statement
This statement does not apply to this article.
Ethics Statement
This research did not involve human participants, animal subjects, or any material that requires ethical approval.
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Clinical Trial Registration
This research does not involve any clinical trials.
Permission to reproduce material from other sources
Not Applicable.
Author Contributions
- Damayanti Dayanand Khutwad: Conceptualization, literature review, and manuscript preparation, including Abstract, Introduction, Plant description, Geographical distribution, collection, and cultivation, Phytoconstituents, pharmacological uses, Adverse effects, Marketed formulations, Homemade remedies, Discussion, and Conclusion
- Kishor Otari: Supervision, critical revision of the manuscript for important intellectual content, and overall guidance.
- Ajay Kale: Literature collection, reference management, manuscript formatting, and proofreading.
References
- Otte C, Gold SM, Penninx BW, et al. Major depressive disorder. Nat Rev Dis Primers. 2016;2:16065. doi:10.1038/nrdp.2016.65.
CrossRef - Malhi GS, Mann JJ. Depression. Lancet. 2018;392(10161):2299-2312. doi:10.1016/S0140-6736(18)31948-2.
CrossRef - Cipriani A, Furukawa TA, Salanti G, et al. Comparative efficacy of antidepressants. Lancet. 2018;391(10128):1357-1366. doi:10.1016/S0140-6736(17)32802-7.
CrossRef - Duman RS, Aghajanian GK. Synaptic dysfunction in depression. Science. 2012;338(6103):68-72. doi:10.1126/science.1222939.
CrossRef - Krishnan V, Nestler EJ. Neurobiology of depression. Nature. 2008;455(7215):894-902. doi:10.1038/nature07455.
CrossRef - Ferrari AJ, Charlson FJ, Norman RE, et al. Global burden of depression. Psychol Med. 2013;43(3):471-481. doi:10.1017/S0033291712001795.
- World Health Organization. Depression Worldwide. Geneva, Switzerland: World Health Organization; 2023.
- Kessler RC, Bromet EJ. Epidemiology of depression. Annu Rev Public Health. 2013;34:119-138. doi:10.1146/annurev-publhealth-031912-114409.
CrossRef - Dantzer R, O’Connor JC, Freund GG, et al. Inflammation and depression. Nat Rev Neurosci. 2008;9:46-56. doi:10.1038/nrn2297.
CrossRef - Miller AH, Raison CL. Inflammatory mechanisms in depression. Nat Rev Immunol. 2016;16(1):22-34. doi:10.1038/nri.2015.5.
CrossRef - Pariante CM. HPA axis and depression. Trends Neurosci. 2017;40(8):464-475. doi:10.1016/j.tins.2017.05.004.
CrossRef - McEwen BS. Stress and brain plasticity. Annu Rev Med. 2017;68:219-233. doi:10.1146/annurev-med-042915-103630.
- Nestler EJ, Hyman SE. Animal models of depression. Nat Neurosci. 2010;13:1161-1169. doi:10.1038/nn.2647.
CrossRef - Willner P. Chronic mild stress model. Neurobiol Stress. 2017;6:78-93. doi:10.1016/j.ynstr.2016.08.002.
CrossRef - Cryan JF, Mombereau C. Behavioral models. Mol Psychiatry. 2004;9:326-357. doi:10.1038/sj.mp.4001454.
CrossRef - Planchez B, Surget A, Belzung C. Animal models of depression: an overview. Dialogues Clin Neurosci. 2019;21(1):41-52. doi:10.31887/DCNS.2019.21.1/bplanchez.
- Steru L, Chermat R, Thierry B, Simon P. Tail suspension test. Psychopharmacology. 1985;85:367-370. doi:10.1007/BF00428203.
CrossRef - Porsolt RD, Le Pichon M, Jalfre M. Forced swim test. Nature. 1977;266:730-732. doi:10.1038/266730a0.
CrossRef - Antkiewicz-Michaluk L. Reserpine model. Pharmacol Rep. 2014;66:163-171. doi:10.1016/j.pharep.2013.10.002.
CrossRef - Hill MN, Hellemans KG, Verma P, et al. Stress models. Neurosci Biobehav Rev. 2012;36:715-731. doi:10.1016/j.neubiorev.2011.10.001.
CrossRef - Sarris J. Herbal medicines in psychiatry. J Affect Disord. 2013;150:1-12. doi:10.1016/j.jad.2013.02.013.
CrossRef - Panossian A, Wikman G. Adaptogens. Pharmacol Ther. 2010;128:1-23. doi:10.1016/j.pharmthera.2010.05.003.
CrossRef - Kumar S. Phytochemicals in CNS disorders. Front Pharmacol. 2017;8:577. doi:10.3389/fphar.2017.00577.
CrossRef - Nabavi SF. Neuroprotective phytochemicals. Neurosci Biobehav Rev. 2015;57:128-147. doi:10.1016/j.neubiorev.2015.08.010.
CrossRef - Lopresti AL, Maes M. Curcumin and depression. J Affect Disord. 2014;167:368-375. doi:10.1016/j.jad.2014.06.001.
CrossRef - Chandrasekhar K, Kapoor J. Ashwagandha trial. Indian J Psychol Med. 2012;34:255-262. doi:10.4103/0253-7176.106022.
CrossRef - Aguiar S, Borowski T. Bacopa monnieri. Evid Based Complement Alternat Med. 2013;2013:717459. doi:10.1155/2013/717459.
CrossRef - Butterweck V. Plant antidepressants. Planta Med. 2003;69:97-109. doi:10.1055/s-2003-37705.
CrossRef - Linde K, Berner MM. St John’s wort. 2005;330:503. doi:10.1136/bmj.330.7490.503.
CrossRef - Tandon VR. Vitex negundo. Indian J Pharmacol. 2005;37:41-44. doi:10.4103/0253-7613.13854.
CrossRef - Rege SD, Geetha T. Antioxidants. Neurochem Int. 2014;78:85-94. doi:10.1016/j.neuint.2014.08.006.
CrossRef - Liu Y. Neuroinflammation. Brain Behav Immun. 2017;66:273-281. doi:10.1016/j.bbi.2017.07.008.
CrossRef - Duman RS, Monteggia LM. Neuroplasticity. Biol Psychiatry. 2006;59:1116-1127. doi:10.1016/j.biopsych.2006.02.013.
CrossRef - Castrén E, Rantamäki T. BDNF. Trends Neurosci. 2010;33:259-267. doi:10.1016/j.tins.2012.12.001.
CrossRef - Black CN, Bot M. Oxidative stress. J Affect Disord. 2015;172:90-97. doi:10.1016/j.jad.2014.09.047.
CrossRef - Sapolsky RM. Stress brain. Arch Gen Psychiatry. 2000;57:925-935. doi:10.1001/archpsyc.57.10.925.
CrossRef - Harmer CJ, Duman RS. Rapid antidepressants. Nat Rev Neurosci. 2017;18:703-713. doi:10.1038/nrn.2017.105.
CrossRef - Wohleb ES. Microglia depression. Nat Rev Neurosci. 2016;17:497-511. doi:10.1038/nrn.2016.69.
CrossRef - Réus GZ. Neurobiology of depression. 2015;300:257-273. doi:10.1016/j.neuroscience.2015.05.038.
CrossRef - Jeon SW, Kim YK. Inflammation and depression are linked. Neurosci Lett. 2017;651:200-205. doi:10.1016/j.neulet.2017.04.056.
CrossRef - Zhang JC. Rapid antidepressant mechanisms. Prog Neurobiol. 2021;198:101905. doi:10.1016/j.pneurobio.2020.101905.
CrossRef - Fakhri S, Pesce M, Patruno A, et al. Attenuation of neuroinflammation in depression by herbal medicines and phytochemicals. 2021;26(17):5196. doi:10.3390/molecules26175196.
CrossRef - Rauf A, Abu-Izneid T, Khalil AA, et al. Berberine as a potential antidepressant: mechanisms of action and therapeutic implications. Biomed Pharmacother. 2021;137:111301. doi:10.1016/j.biopha.2021.111301.
CrossRef - Nabavi SF, Sureda A, Daglia M, et al. Curcumin and depression: a review of clinical and preclinical studies. Phytother Res. 2015;29(1):17-25. doi:10.1002/ptr.5214.
CrossRef - Sarris J, Panossian A, Schweitzer I, Stough C, Scholey A. Herbal medicine for depression, anxiety and insomnia: a review of psychopharmacology and clinical evidence. Eur Neuropsychopharmacol. 2011;21(12):841-860. doi:10.1016/j.euroneuro.2011.04.002.
CrossRef
Abbreviations
BDNF: Brain-Derived Neurotrophic Factor
CA1: Cornu Ammonis Area 1 (Hippocampus)
CA3: Cornu Ammonis Area 3 (Hippocampus)
CNS: Central Nervous System
CUMS: Chronic Unpredictable Mild Stress
DA: Dopamine
FST: Forced Swim Test
GABA: Gamma-Aminobutyric Acid
HPA Axis: Hypothalamic–Pituitary–Adrenal Axis
IL-1β: Interleukin-1 Beta
IL-6: Interleukin-6
MAO: Monoamine Oxidase
MAOIs: Monoamine Oxidase Inhibitors
MDD: Major Depressive Disorder
NE: Norepinephrine
ROS: Reactive Oxygen Species
SSRIs: Selective Serotonin Reuptake Inhibitors
TCA: Tricyclic Antidepressants
TNF-α: Tumor Necrosis Factor-Alpha
TST: Tail Suspension Test
Accepted on: 22-05-2026
Second Review by: Dr. Ramya Sri and Dr. Shankar Yelmame
Final Approval by: Dr. Mohammad Fareed








![]()
![]()
