

Manuscript accepted on : 12-June-2019
Published online on: 29-06-2019
Plagiarism Check: Yes
Reviewed by: Osama Matloup
Second Review by: Tarek Morsy
Seyed Masoud Davoodi1, Mohsen Danesh Mesgaran*1, Ali Reza Vakili1, Reza Valizadeh1 and Abdollah Ghasemi Pirbalouti2
1Department of Animal Science, Faculty of Agriculture, Ferdowsi University of Mashhad, P.O. Box 91775-1163, Mashhad, Iran.
2Department of Medicinial Plants, Faculty of Agriculture, Shahrekord Branch, Islamic Azad University, P.O. Box 166, Shahrekord, Iran.
Corresponding Author E-mail: danesh@um.ac.ir
DOI : http://dx.doi.org/10.13005/bbra/2749

ABSTRACT: Present study was conducted to investigate the effect of including plant essential oils on in vitro ruminal fermentation and microbial nitrogen synthesis of a dairy cow diet rich in concentrate. The treatments consisted of the diet alone (control; BD) as well as containing 50 and 100 μl L-1 essential oil of thyme (BDT), mint (BDM), savory (BDS), or a mixture of the essential oils at the rate of 1:1:1 (BDmix). Essential oils decreased gas production at 24, 48 and 96 h of incubation compared with that of BD. However, mint at the rate of 50 or 100 μl L-1 resulted an increase in the microbial nitrogen when compared to BD, BDS and BDT. The nitrogen content of truly undegraded residu (NDFN) content and NH3-N concentration were lower, while the dry matter digestibility was greater in the BDmix, regardless of dosage levels, as compared with the control. The inclusion of a mixture of essential oils at 50 μl L-1 to the basal diet caused intensified dry matter disappearance, in comparison to other treatments. Results showed that the synergetic effects of essential oils together in a dairy cow diet of rich in concentrate can alter rumen microbial fermentation and improve microbial protein yield.
KEYWORDS: Essential Oils; In Vitro Fermentation; Mint; Microbial Nitrogen; Savory; Thyme
Download this article as:| Copy the following to cite this article: Davoodi S. M, Mesgaran M. D, Vakili A. R, Valizadeh R, Pirbalouti A. G. In vitro Effect of Essential Oils on Rumen Fermentation and Microbial Nitrogen Yield of High Concentrate Dairy Cow Diet. Biosci Biotech Res Asia 2019;16(2). |
| Copy the following to cite this URL: Davoodi S. M, Mesgaran M. D, Vakili A. R, Valizadeh R, Pirbalouti A. G. In vitro Effect of Essential Oils on Rumen Fermentation and Microbial Nitrogen Yield of High Concentrate Dairy Cow Diet. Biosci Biotech Res Asia 2019;16(2). Available from: https://bit.ly/2S6hyv0 |
Introduction
In recent years, antibiotics and antimicrobial compounds produced by microorganisms have played an important role in animal diets to increase their performances and decrease mortality (Benchaar et al., 2008). However, public concern over routine use of antibiotics in animal nutrition has increased due to the potential development of antibiotic resistant in animals and humans (Benchaar et al., 2006). As a result, alternative feed additives such as aromatic plants and their extracts were researched instead of antibiotics and they have gained interest as growth and health promoters (Benchaar et al., 2006). Essential Oils (EOs) are volatile, lipophilic substances obtained from plant materials which are gained generally by steam distillation mechanical separation or solvent extraction (Wallace, 2002; Castillejos et al., 2007; Hart et al., 2008). Hart et al., (2008), described that the concentration of EOs may vary depending on the stage of plant growth, different part of plant, amount of moisture received, light, and temperature. Essential oils are complex mixtures of secondary plant metabolites that demonstrate a broad spectrum of antimicrobial properties, particularly against gram positive bacteria (Burt, 2004). They are naturally occurring plant components and are considered safe for human and animal consumption (FDA, 2003; Benchaar et al., 2006). These appear to be selective in their antifungal, antiviral, bactericidal, and bacteriostatic effects on microorganisms (Burt, 2004; Janssen et al., 1986). Oh et al., (1967) and then Nagy and Tengerdy (1967), were the first to investigate effects of EOs on in vitro ruminal microbial fermentation and observed that EOs inhibited gas production, however the degree of inhibition was dependent on the chemical structure of the EOs.
The research of Smith-Palmer et al., (1998), determined that out of their EOs test articles, the oils of cinnamon and thyme exhibited the greatest inhibitory effects with bacteriostatic activity at a concentration of 0.075% or less against Escherichia coli, Staphylococcus aureus, Listeria monocytogenes, Salmonella enteritidis, and Campylobacter jejuni. However, negative gram organisms in this study (and others) were slightly less susceptible to EOs than positive gram bacteria (Smith-Palmer et al., 1998; Burt, 2004). Certain phenolic compounds such as thymol (thyme) could inhibit negative gram bacteria as well as positive gram bacteria. The mechanism by which most EOs are thought to exert their antibacterial effects is by disrupting cell wall structures due to their lipophilic character. The chemical components of EOs interrupt electron transport, ion gradients, protein translocation, phosphorylation steps, and other enzyme-dependent steps, causing the bacterium to lose chemi-osmotic control (Ultee and Kets, 1999; Cox et al., 2000; Dorman and Deans, 2000) . A number of researches were induced by the well-documented antimicrobial effectiveness of EOs improve their feed efficiency and so lead to an increased production efficiency animal (Wallace, 2002). During the past few years, various studies have been conducted to determine the effects of herbal plant EOs on rumen microbial fermentation and dry matter disappearance (Hart et al., 2008; Busquet et al., 2006). It seems that a level of EOs used in the diet and the source of supplied them from herbal plants to be important as regards its positive effects on rumen microbial fermentation.
Essential oils provide a valuable approach to manipulate ruminal fermentation to decrease methane and NH3-N concentration and increased the amount of microbial protein production (MP) by ruminant (Patra and Saxena, 2010; Menke and Steingass, 1988). Microbial protein production plays a pivotal role in ruminant nutrition (Srinivas and Krishnamoorthy, 2013). Various studies have observed contradictory results when improving microbial protein (MP) by the inclusion of essential oil (EOs). Menke and Steingass (Menke and Steingass, 1988), noted that plant extracts increase the degradation of cell wall constituents, yield and efficiency of MP synthesis. However, Kumar et al., (2011), observed no adverse impact of EOs on MP of buffaloes, with this conflict being possibly related to inherent detoxification ability of bacteria or neglecting the dosage and type of EOs. Furthermore, the stereochemical properties of EOs can vary and depend on the method of extraction (Scorzoni et al., 2007). However, extraction products may also vary qualitatively and quantitatively in their composition (Angioni et al., 2006). Therefore, the objective of the present experiment was to evaluate the in vitro effects of various essential oils on rumen fermentation and microbial nitrogen synthesis of a dairy cow diet rich in concentrate.
Materials and Methodology
Herbal plants were collected from botanical garden of chaharmahall and bakhtiari province (shahrekord city, Iran). The essential oil content for each plant was obtained through steam distillation by Clevenger Method. A basal experimental diet (BD) used for batch cultures was a dairy cow diet rich in concentrate. Diet ingredients and chemical composition are presented in Table 1.
Table 1: Ingredients (% DM) and nutrient composition of basal diet.
| Ingredients | % |
| Corn silage | 20 |
| Alfalfa hay | 20 |
| Corn grain | 14 |
| Barley grain | 17.5 |
| Soybean meal | 11 |
| Wheat bran | 10 |
| Canola meal | 7 |
| Mineral and vitamin premix1 | 0.5 |
| Chemical composition (% DM) | |
| Crude protein | 17.2 |
| Neutral detergent fiber | 32.1 |
| Acid detergent fiber | 19.2 |
| Ether extract, | 2.1 |
| Non-fiber carbohydrate2 | 44. 4 |
1Supplied 195 g/kg of Ca, 21 g/kg of Mg, 2.2 g/kg of Mn, 0.3 g/kg of Zn, 0.3 g/kg of Cu, 0.12 g/kg of I, 0.1 g/kg of Co, 600,000 IU/kg of vitamin A, 200,000 IU/kg of vitamin D, and 0.2 g/kg of vitamin E, 0.025 g/ kg of Se.
2Calculated by difference 100 − (% NDF + % CP + % fat + % ash).
The treatments included BD (control), BD + thyme (BDT), BD + mint (BDM), BD + savory (BDS) and BD plus a mixture of thyme, mint and savory at the rate of 1:1:1 (BDmix), 6 replicates per each treatment of three runs. Two different doses were considered for each essential oil as 50 and 100 μl L-1 of the total culture medium. The gas production procedure and microbial N assay were performed as described by Grings et al., (2005). Rumen inoculum was collected from three rumen fistulae Holstein lactating dairy cows (620 ± 10 kg of BW, 310 ± 8 DIM, mean ± SD) and rumen liquor was handled under a constant stream of CO2, all containers used were pre-warmed at 39ºC and filled with CO2. The cows were offered a diet (g/kg DM) containing alfalfa hay (181), corn silage (289), wheat straw (31), barley grain (165), corn grain (114), soybean meal (124), wheat bran (93), vitamin and mineral premix (10). Ruminal content was immediately transferred to the laboratory, then blended and strained through four layers of cheesecloth to eliminate large feed particles and transferred to the laboratory in a pre-warmed thermos.A sample of 250 mg was weighed into a 125-ml serum bottles in 3 runs with 6 replicates.
The filtrate was then mixed with carbonate buffer (containing ammonium bicarbonate at 4g/l) and sodium bicarbonate (35g/l in N-rich incubation medium and only sodium bicarbonate at 39.25g/l in N-low medium), macro-mineral solution (5.7g anhydrous Na2HPO4, 6.2g anhydrous KH2PO4 and 0.6g MgSO4·7H2O per liter), and deionized water in a ratio of 1:1:0.5:1.5 and 0.1 ml micro-mineral solution (13.2g CaCl2·2H2O, 10.0g MnCl2·4H2O, 1g CoCl2·6H2O and 8.0 g FeCl3·6H2O per 100ml) was added. The medium was then reduced by addition of 41.7ml reducing agent (40ml deionized water, 1ml 1N NaOH and 1g Na2S·9H2O) per liter. Twenty milliliters of medium were dispensed into a 125ml glass serum bottle whose top were stopped with rubber and aluminum cabs and placed in a water bath for 96 h at 39ºC. Blank samples were also incubated simultaneously to make correction in gas production, if any, from the medium. Gas production was measured at 2, 4, 6, 8, 10, 12, 24, 48, 72 and 96 h of the incubation by inserting a 23 gauge (0.6mm) needle attached to a pressure transducer (model PX4200-015GI, Omega Engineering, Inc. Laval, Que., Canada) connected to a visual display (Data Track, Christchurch, UK) into the head space of serum bottles.The transducer was then removed leaving the needle in place to permit venting. Pressure values were corrected for the amount of substrate organic matter (OM) incubated and the gas released from negative controls.
In order to prevent accumulation of produced gases, the gas in the head space of each bottle was released. After subtraction of gas production from blank bottles, data were fitted to the following exponential model, based on Orskov and McDonald, (1979) model without lag phase: G = b (1 − e−ct), where G is the cumulative gas volume produced at time t, b is the asymptotic (or maximal) gas volume, i.e., G(t → ∞), and c is the gas production rate.Then, half time of maximal gas production (t1/2) was calculated and same incubation with similar dietary substrates was done to assess the microbial N production. Microbial N production at t1/2 was indirectly estimated by differentially quantifying all N sources, except microbial N, and includes use of information on the N content of diet, truly un-degraded N (NDFN) in diet samples at t1/2, and the change in NH3–N levels in the incubation medium between 0 hour of incubation and t1/2 using the equations: Microbial N production at t1/2 = dietary N + ∆NH3–N – NDFN at t1/2.Where ∆NH3–N=NH3–N in 0 hour blanks − NH3–N in diet incubations at t1/2. Dry matter degradability of samples at t1/2 was measured by refluxing the incubation residue with neutral detergent (ND) solution for 1 hour.
Analytical Methods
Dietary samples were oven dried at 60 ºC for 48h, then ground to pass through a 1-mm screen in a Wiley mill (standard model 4; Arthur H. Thomas Co., Philadelphia, PA) before chemical analyses. The measurement of diet nitrogen was conducted by Kjeldahl analysis (AOAC,1990). Crude protein was determined as nitrogen × 6.25. The fat content of the diet was determined using a Soxtec system HT6 apparatus according to AOAC, (1990). The concentration of NDF and ADF was determined as described by Van Soest et al., (1991) without the use of sodium sulfite and heat-stable α-amylase. The NFC content of the diets was calculated as 1000 − (NDF + CP + ether extract + ash).
Statistical Analysis
In order to assess the differences between dietary treatments, the data were analyzed in a completely randomized design using the general linear models procedure of SAS, (2003). In this analysis, mean values of various parameters of each diet (e.g. microbial N) were compared using the Duncan’s multiple range tests. Contrasts were tested using the CONTRAST statement of SAS. Standard errors of means were calculated from the residual mean square in the analysis of variance. Standard errors of means were calculated from the residual mean square in analysis of variance. Significance was declared at P≤ 0.05.
Results
In Vitro Gas Production
As illustrated in Table 2, the inclusion of EOs on diet, regardless of dosage levels, had no significant effect on asymptotic gas production (b), first-order fractional rate constant (c), and the time (h), when half of the asymptotic gas volume was produced (t1/2). The gas volume at t1/2 was the highest in BD and was the same across BDT, BDS, BDM and BDmix at both 50 and 100 µl levels. Furthermore, gas production at 24, 48 and 96 h decreased with supplementing EOs, regardless of dosage levels and sort of plant EOs, as compared with that of control. However, the increase of each essential oil level from 50 µl to 100 µl did not have a significant effect on reduction of gas volume. Nevertheless, 50 µl of each essential oil had a minimal inhibiltory effect on gas production.
Table 2: Effect of including plant essential oils (applied at the rate of 50 or 100 μl/L of medium) in a high concentrate diet on asymptotic gas production (b), first-order fractional rate constant (c), gas production at 24, 48, 96 h and t1/2.
| Treatments1 | ||||||||||||
| BD | BDT | BDM | BDS | BDmix | P value | |||||||
| Parameters | Concentrate of EO added to basal diet | Concentrate of EO added to basal diet | Concentrate of EO added to basal diet | Concentrate of EO added to basal diet | SEM | BD vs. | BD+50 μl/L EO vs. BD+100 μl/L EO | |||||
| (μl l-1) | (μl l-1) | (μl l-1) | (μl l-1) | BD + EOs | ||||||||
| 50 | 100 | 50 | 100 | 50 | 100 | 50 | 100 | |||||
| b | 54.11 | 56.03 | 54.46 | 51.21 | 51.1 | 54.37 | 52.4 | 56.53 | 55.97 | 1.9 | 0.37 | 0.51 |
| c | 0.08 | 0.07 | 0.08 | 0.07 | 0.07 | 0.07 | 0.07 | 0.08 | 0.07 | 0.003 | 0.62 | 0.69 |
| T1/2 | 2.3 | 2.56 | 2.1 | 3.01 | 2.41 | 2.9 | 3.02 | 3.28 | 3.44 | 0.51 | 0.26 | 0.48 |
| Gast1/2 | 20.10a | 16.94b | 17.60b | 17.33b | 18.02b | 17.10b | 17.51b | 16.99b | 17.88b | 1.09 | 0.001 | 0.53 |
| Gas24 | 49.70a | 21.12b | 19.29b | 20.18b | 25.05b | 28.66b | 19.37b | 18.19b | 20.24b | 4.2 | 0.001 | 0.47 |
| Gas48 | 54.60a | 39.55b | 38.38b | 39.00b | 41.12b | 40.99b | 39.12b | 39.77b | 43.16b | 2.9 | 0.001 | 0.49 |
| Gas96 | 77.39a | 67.40b | 65.12b | 68.23b | 69.44b | 69.20b | 66.23b | 68.74b | 70.40b | 2.04 | 0.001 | 0.53 |
1control (BD), BD + thyme (BDT), BD + mint (BDM), BD + savory (BDS) and BD plus a mixture of thyme, mint and savory at the rate of 1:1:1 (BDmix)
Microbial Nitrogen Production
Data of Microbial N yield (MN, mg), Nitrogen content of truly undegraded residu (NDFN), NH3-N concentrate, dry matter digestibility,undigested nitrogen (UDN, mg) and the ratio of MN to DMD at t1/2 are presented in Table 3. Microbial nitrogen was the highest in BD mixed with EOs, while it was the lowest in BD without any essential oil supplementation. However, no detectable differences were observed between the two dosages (50 and 100 µl) of EOs among treatments for microbial nitrogen yield (P>0.05). An increase in microbial nitrogen yield (MN) in the diet supplemented with a mixture of EOs is undoubtedly due to the synergetic effect of EOs when used together. The NDFN content in BDmix was lower than that of BD plus thyme, mint and savory. Like wise, diets containing 100 µl of EOs had a lower NDFN than diets receiving 50 µl. The inclusion of essential oils to BD resulted in diminished NH3-N concentration as compared with the control. The BD plus mint essential oil, regardless of its level, had a lower NH3-N concentration than BD plus thyme and savory essential oil.
Table 3: Effect of including plant essential oils (applied at the rate of 50 or 100 μl/L of medium) on Microbial N yield (MN), NDFN, NH3-N, DMD (%), the ratio of microbial nitrogen to dry matter digestibility (MN/DMD), and undigested nitrogen (UDN) at t1/2.
| Treatments1 | ||||||||||||
| BD | BDT | BDM | BDS | BDmix | P value | |||||||
| Parameters | Concentrate of EO added to basal diet | Concentrate of EO added to basal diet | Concentrate of EO added to basal diet | Concentrate of EO added to basal diet | SEM | BD vs. BD + EOs | BD+50 μl/L EO vs. BD+100 μl/L EO | |||||
| (μl l-1) | (μl l-1) | (μl l-1) | (μl l-1) | |||||||||
| 50 | 100 | 50 | 100 | 50 | 100 | 50 | 100 | |||||
| MN (mg) | 4.10e | 4.31cde | 4.28cde | 4.40bc | 4.38bc | 4.28cde | 4.16cde | 4.57a | 4.64a | 0.09 | 0.01 | 0.39 |
| NDFN (mg) | 1.70d | 1.91b | 1.67f | 2.00a | 1.99a | 1.83c | 1.75e | 1.67g | 1.68f | 0.02 | 0.01 | 0.02 |
| NH3-N | 1.43a | 0.88bc | 1.04b | 0.71c | 0.74c | 1.04b | 1.03b | 0.86bc | 0.74c | 0.01 | 0.01 | 0.04 |
| DMD % | 37.16d | 41.88bc | 42.11bc | 42.55bc | 42.67bc | 43.17bc | 41.38c | 45.48a | 45.22ab | 1.07 | 0.01 | 0.21 |
| DMD (mg) diss.) | 92.91d | 104.72bc | 105.28bc | 106.38bc | 106.69bc | 107.94bc | 103.47c | 113.72a | 110.55ab | 2.69 | 0.01 | 0.36 |
| UDN (mg) | 1.53ab | 1.61ab | 1.48ab | 1.62ab | 1.68a | 1.48ab | 1.61ab | 1.38b | 1.43b | 0.1 | 0.03 | 0.34 |
| MN/DMD | 0.044 | 0.041 | 0.041 | 0.041 | 0.042 | 0.04 | 0.039 | 0.041 | 0.04 | 0.02 | 0.39 | 0.32 |
Disscusion
The BD diet resulted in the greatest gas production as compared to other treatments. These findings might due to the inhibitory effect EOs on methane production, which is in line with Patra and Yu, (2012) findings who noted that total gas and methane production by the ruminal cultures decreased with inclusion of EOs in diet. Several studies have documented a reduction in methane production by EOs (Agarwal et al., 2009; Chaves et al., 2008). Macheboeuf et al., (2008) reported that decrease in gas production up to 83% after addition of oregano to the incubation media was occured. Carvacerol and thymol caused a reduction in gas production (Benchaar et al., 2007). Garcia-Gonzalez et al., (2008) also indivated that the plant active compounds can reduce gas production through lessening methane production by affecting protozoa population. Methane production decreased in a batch culture when essential oils were added at 1 μL/mL or at 70, 140 and 280 ppm (Jahani-Azizabadi et al., 2014; Jahani-Azizabadi et al., 2011). However, the in-vivo study of Beauchemin and McGinn, (2006) did not reveal any effect on methanogensis. The inconsistencies among results obtained in different studies may be attributed to the doses used, the main active compounds in plants, and the basal diet.
The decrease in NH3-N concentration with the addition of EOs was consistent with inhibition proteolysis, peptidolitic and deamination process of rumrn microorganisms (Castillejos et al., 2004). McIntosh et al., (2003) demonstrated that a commercial blend of essential oil reduced the rate of amino acid deamination and inhibited the growth of a specific group of ammonia bacteria. Brochers, (1965) and Castillejos et al., (2007) observed that an addition of thyme and oregano EOs to a medium containing rumen liquid resulted in an accumulation of amino acids nitrogen and a decrease in the ammonia nitrogen. Wallace et al., (2002) noted that the antimicrobial properties of EOs affected rumen microbial activities by reducing the dietary protein degradation and deamination, thereby enhancing rumen N escape. On the other hand, dry matter digestibility was influenced by incorporation of EOs, where the effect was the greatest using a mixture of essential oils at both dosages, while it was the lowest in the control diet. We speculate that the lowest dry matter digestibility in the control diet was congruent with greater NDFN content. Undigested nitrogen content was lower in the diet containing a mixture of essential oils at both levels. The ratio of microbial nitrogen and total gas production to dry matter disappearance was the highest in BD when compared to BDmix (Fig. 1).
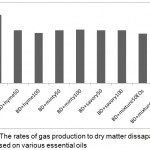 |
Figure 1: The rates of gas production to dry matter dissaparance of diet based on various essential oils.
|
Conclusion
Overall, the present results suggest that the plant essential oils used, especially when mixed EOs were used, resulted in a considerable effect on rumen microbial fermentation and microbial protein yield. Results obtained from the present study demonstrated that by using these EOs in ruminant, we may manipulate rumen fermentation rate by changing in microbial protein yield.
Acknowledgements
This work was conducted with the financial support of the Ferdowsi University of Mashhad, Iran.
References
- Benchaar C., Calsamiglia S., Chaves A. V., Fraser G. R., Colombatto D. A review of plant-derived essential oils in ruminant nutrition and production. J. Anim. Feed Sci. Technol. 2008;145:209-228.
- Benchaar C., Duynisveld J. L., Charmley E. Effects of monensin and increasing dose levels of a mixture of essential oil compounds on intake, digestion and growth performance of beef cattle. Can. J. Anim. Sci. 2006;86:91-96.
- Wallace R. J. Antimicrobial properties of plant secondary metabolites. Proc. Nutr. Soc. 2002;63:621-629.
- Castillejos L., Calsamiglia S., Ferret A., Losa R. Effects of dose and adaptation time of a specific blend of essential oil compounds on rumen fermentation. J. Anim. Feed Sci. Technol. 2007;132:186-201.
- Hart K. J., Yanez-Ruiz D. R., Duval S. M., McEwan N. R., Newbold C. J. Plant extracts to manipulate rumen fermentation. Anim Feed Sci Technol 2008;147: 8-35.
- Burt S. Essential oils: Their antibacterial properties and potential applicants in foods-a review.Int. J. Food Microbiol. 2004;94:223-253.
- FDA. Freedom of information summary.Original new drug application NADA 141-221.Ractopamine hydrochloride (OPTAFLEXX 45). Type A medicated article for beef cattle. 2003.
- Janssen A. M., Chin N. L., Scheffer J. J. M., Baerheim-Svendsen A. Screening for antimicrobial activity of some essential oil by the ager overley technique. Pharm Weekblad Sci. 1986;8:289-292.
- Oh H. K., Sakai T., Jones M. B., Longhurst W. M. Effect of various essential oils isolated from douglasfir needles upon sheep and deer rumen microbial activity. J. Appl. Microbiol. 1967;15:777-784.
- Nagy J. G., Tengerdy R. P. Antibacterial Action of Essential Oils of Artemisia as an Ecological Factor: I. Antibacterial Action of the Volatile Oils of Artemisia tridentata and Artemisia nova on Aerobic Bacteria. J. Appl. Microbiol. 1967;15:819-821.
- Smith-Palmer A., Stewart J., Fyfe L. Antimicrobial properties of plant essential oils and essences against five important food-borne pathogens.Lett. J. Appl. Microbiol. 1998;26:118-122.
- Ultee A., Kets E. P. W. Mechanisms of action of carvacrol on the food-borne pathogen Bacillus cereus. J. Appl. EnviroMm Microbiol. 1999; 65:4606-4610.
- Cox S. D., Mann C. M., Markham J. L., Bell H. C., Gustafson J. E. The mode of antimicrobial action of the essential oil of Melaleucaalternifolia (tea tree oil). J. Appl. Microbiol. 2000;88:170-175.
- Dorman H. J. D., Deans S. G. Antimicrobial agents from plants: antimicrobial activity of plant volatile oils. J Appl Microbiol 2000;88:308-316.
- Busquet M., Calsamiglia S., Ferret A., Kamel C. Plant extracts affect in vitro rumen microbial fermentation. J. Dairy Sci. 2006;89:761-771.
- Patra A. K., Saxena J. A New perspective on the use of plant secondary metabolites to inhibit methanogenesis in the rumen. J. Phytochem. 2010;71:1198-1222.
- Menke K. H., Steingass H. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim Res Develop 1988;28:7-55.
- Srinivas B., Krishnamoorthy U. Panoply of microbial protein production in ruminants–A review.The Indian J. Anim. Sci. 2013;83:331-346.
- Kumar S., Wahab N., Warikoo R. Bioefficacy of Menthapiperita essential oil against dengue fever mosquito Aedesaegypti L. Asian Pacific J. tropical biomedicine 2011;1:85-88.
- Scorzoni L., Benaducci T., Almeida A. M. F., Silva D. H. S., Bolzani V. D. S. The use of standard methodology for determination of antifungal activity of natural products against medical yeasts Candida sp and Cryptococcus sp. Brazilian J. Microbiol. 2007;38:391–397.
- Angioni A., Barra A., Coroneo V., Dessi S., Cabras P. Chemical composition, seasonal variability, and antifungal activity of Lavandula stoechas L. ssp. stoechas essential oils from stem/leaves and flowers. J. Agricult. and Food Chem. 2006;54:4364–4370.
- Grings E. E., Blümmel M., Südekum K. H. Methodological considerations in using gas production techniques for estimating ruminal microbial efficiencies for silage-based diets. J. Anim. Feed Sci. Technol. 2005;123:527-545.
- Orskov E. R., McDonald I. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agricult. Sci. 1979;92:499-503.
- AOAC. Method 990-03. Official Methods of Analysis, First Supplement. Arlington, VA, USA, 1990.746 pages.
- Van Soest P. J., Robertson J. B., Lewis B. A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991;74:3583-3597.
- SAS. Statistical Analysis System. User’s Guide: Statistics,Version 9.2. SAS Institute, Cary, NC, USA.2003.
- Patra A. K., Yu Z. Effects of essential oils on methane production and fermentation by, and abundance and diversity of, rumen microbial populations. J. Appl. Environ. Microbiol. 2012;78:4271-4280.
- Agarwal N., Shekhar C., Kumar R., Chaudhary L. C., Kamra D. N. Effect of peppermint (Menthapiperita) oil on in vitro methanogenesis and fermentation of feed with buffalo rumen liquor. J. Anim. Feed Sci. Technol. 2009;148:321-327.
- Chaves A. V., Stanford K., Dugan M. E. R., Gibson L. L., McAllister T. A. Effects of cinnamaldehyde, garlic and juniper berry essential oils on rumen fermentation, blood metabolites, growth performance, and carcass characteristics of growing lambs. J. Liv. Sci. 2008;117:215-224.
- Macheboeuf D., Morgavi D. P., Papon Y., Mousset J. L., Arturo-Schaan M. Dose-response effects of essential oils on in vitro fermentation activity of the rumen microbial population. J. Anim. Feed Sci. Technol. 2008;145:335-350.
- Benchaar C., Petit H. V., Berthiaume R., Ouellet D. R., Chiquette J. Effects of Essential Oils on Digestion, Ruminal Fermentation, Rumen Microbial Populations, Milk Production, and Milk Composition in Dairy Cows Fed Alfalfa Silage or Corn Silage. J. Dairy Sci. 2007;90:886-897.
- Garcia-González R., Lopez S., Fernandez M., Gonzalez J. S. Dose response effects of Rheum officinale root and Frangula alnus bark on ruminal methane production in vitro. J. Anim. Feed Sci. Technol. 2008;145:319-334.
- Jahani-Azizabadi H., Danesh Mesgaran M., Vakili A., Rezayazdi K. Effect of some plant essential oils on in vitro ruminal methane production and on fermentation characteristics of a mid-forage diet. J. Agric. Sci. Technol. 2014;16:1543-1554.
- Jahani-Azizabadi H., Danesh Mesgaran M., Vakili A., Rezayazdi K., Hashemi M. Effect of various medicinal plant essential oils obtained from semi-arid climate on rumen fermentation characteristics of a high forage diet using in vitro batch culture. African J. Microbiol. Res. 2011;5:4812-4819.
- Beauchemin K. A., McGinn S. M. Methane emissions from beef cattle: Effects of fumaric acid, essential oil, and canola oil. J. Anim. Sci. 2006;84:1489-1496.
- McIntosh F. M., Williams P., Losa R., Wallace R. J., Beever D. A. Effects of essential oils on ruminal microorganisms and their protein metabolism. J. Appl. Environ. Microbial. 2003;69:5011-5014.
- Brochers R. Proteolytic activity of rumen fluid in vitro. J. Anim. Sci. 1956;24:1033–1038.

This work is licensed under a Creative Commons Attribution 4.0 International License.



