

Manuscript accepted on : September 07, 2011
Published online on: --
Percy Maruwa Chimwamurombe
Department of Biological Sciences, University of Namibia, P. Bag 13301, Windhoek Namibia.
Corresponding Author E-mail: pchimwa@unam.na
ABSTRACT: Marama bean [Tylosema esculentum (Burchell) Schreiber] is indigenous to the dry parts of Southern Africa. It is a staple food for the Khoisan and Bantu people from those areas. In Namibia it grows wild mainly in Omaheke and Otjizondjupa regions. Marama, which has not been domesticated, has a large tuber and pods containing 1-2 oil and protein-rich seeds with a nutritional value similar to soybean. Protein content is 30-39% (similar to soy bean) and oil content is 30-43%. In addition it produces a edible tuber tht is rich in starch. Unfortunately, harvesting in the wild is very extensive and random by local people with such intensity that some genotypes of marama bean are becoming endangered with extinction or have already become extinct. This article reviews the current body literature on marama bean, domestication of selected major legumes and lastly reports on the progress made in the initial attempts to domesticate this plant by local communities in Namibia. The process starts off selecting marama genotypes with superior traits. These traits include early germination and number of seeds per pod. The University of Namibia has been developing marama for its introduction as a crop alternative for dry lands of Namibia and the Kalahari sandy regions of Southern Africa and also as a response to climate change effects where crops such as maize and millets have been failing due to alternating floods and drought. Marama has a huge potential to address the problem of malnutrition and hunger in the arid areas of Southern Africa, especially knowing that under climate change Africa will become drier and drier.
KEYWORDS: Tylosema esculentuml; Breeding; Conservation genetics
Download this article as:| Copy the following to cite this article: Chimwamurombe P. M. Domestication of [Tylosema esculentum (Burchell) Schreiber] (Marama bean): A work in Progress in Namibia. Biosci Biotech Res Asia 2011;8(2) |
| Copy the following to cite this URL: Chimwamurombe P. M. Domestication of [Tylosema esculentum (Burchell) Schreiber] (Marama bean): A work in Progress in Namibia. Biosci Biotech Res Asia 2011;8(2). Available from: https://www.biotech-asia.org/?p=9420/ |
Introduction
Domestication leads to increased adaptation of plants and animals to cultivation or rearing and utilization by humans (Gerts, 2004). Domestication of crops and animals is widely taken for granted. However, the global climate change coupled with increasing demand for food against decreasing availability land for agriculture has led to a new approach to meet food and feed needs of the world. All these and other factors have culminated into the need to diversify crop agriculture. Generally, the process of plant domestication of takes place over a very long period of time during which people will be selecting for special traits on the particular plant. As this process takes place, the plant inevitably will also be losing its ability to survive on its own in the wild without help or interference from the domesticator (Hymowitz, 1970). Common traits that mankind domesticate for include amongst others, high yield, good taste, pest and disease tolerance and of late flood, heat and late drought tolerance (Gerts, 2004). New technologies have been developed to make domestication happen in less than the normal time. A typical plant domestication process is a result of a carefully chosen interplay between the environment, man and the plant itself (Figure 1). There are key requirements for each of the three factors at play. These are environmental demands like water and temperature, humankind demands like high yield, good taste and knowledge of the biology of the plant itself (Gerts, 2004).
Regarding the plant it is important to understand the basic scientific facts about the growth characteristics of the plant. This informs on what sort of plant the domesticator requires. Understanding what possible response the plant can give to which environmental conditions, human needs and activities also play a key role in informing what trait of the plant to domesticate for. Morden science now allows the three factors to be studied concurrently thus helping to completely domesticate a plant in relatively shorter time period.
In this article, a critical evaluation is going to be made on the domestication of a few food legumes, and conclude by highlighting current efforts in the domestication of Tylosema esculentum (Marama bean) in Namibia. Marama bean is currently an edible wild, perennial, non-nitrogen fixing and drought avoiding legume that in native to the deep Kalahari sandy and arid regions of Southern Africa. It is rich in proteins and oils and competes with most legumes or even surpasses them in nutrient content before even before any dedicated improvements have been done on it yet. The root tuber is a rich source of water and starch.
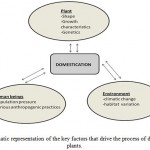 |
Figure 1: Schematic representation of the key factors that drive the process of domestication of plants.
|
Description about domestication process of some major food legumes
Food legumes have long been associated with mankind for several millennia ago. Understandably so because of the nutrient attributes that they contribute to human diet. Of specific importance are the proteins, fatty acids and starch that are contained in the seeds of the legumes. The legumes have also been popular for their medicinal uses among the native folk of many parts of the world, even though this came as an additional gratuitous revelation to the characteristics of the legumes (Hymowitz, 1970).
Major food legumes include, Phaseolus vulgaris (the common bean), Glycine max (soy bean), Vigna subterranea (bambara ground nuts), pea (Pisum sativum), chickpea (Cicer arietinum), broad bean (Vicia faba), pigeon pea (Cajanus cajan), cowpea (Vigna unguiculata), peanut (Arachis hypogeae). Some of the legumes are annuals and others are perennial. The annual legumes are more popular and preferred because they are quicker to reach the market. The domestication process of these major legumes has started from various parts of the world depending on the centre of origin of the legume and has also taken different periods of time. For example soy bean was domesticated in North China since the 11th century BC (Hymowitz, 1970). For human consumption (and all monogastric consumers) soybean must be cooked because it is toxic when eaten raw due to anti-nutritional trypsin and inhibitors that it contains. For cowpea the information regarding its time of domestication is unclear, however the centres of diversity of in West Africa and North-East Africa, even though as early as 1500BC cowpea had started spreading to India (Coulibaly et al., , 2002). The cowpeas must be cooked for human consumption, otherwise if eaten raw they may have fatal results on the consumer. From these two examples, it is clear that even though domestication and carefully selection has taken place for a long time in the past, there are still some attributes that the two plants, soybean and cow peas and indeed most food legumes still have that make them toxic if they are not carefully processed for consumption. Remarkable advances have been in various food legumes so much that they there are many cultivars of the various food legumes. These can be grown annually at different seasons, different desired time periods of growth and suiting various markets. It must still be keep at the back of our minds that even those these major food legumes have been domesticated since a long time ago, the selection and breeding for other interesting characteristics is still going on and even being fast tracked by genetic modification to introduce those traits that otherwise would be impossible from a genetic perspective using natural reproductive processes.
Emphasis on specific legume traits
Legumes have a variety of traits that are typical to them. The term legume is a usual reference to all the plants that bear seeds in a pod. However, the traits of the leguminous plants allow for further taxonomic classification of this broad group. Although Fabaceae is the name used for the family by many, others consider its use to be ambiguous. Therefore this very large family of 650 genera and over 18,000 species is split into three subfamilies or sometimes three separate families (Castro et al, 2005):
– Subfamily Caesalpinioideae or family Caesalpiniaceae
– Subfamily Minosoideae or family Mimosaceae
– Subfamily Faboideae (= Papilionoideae) or family Papilionaceae (= Fabaceae sensu stricto).
Thus Fabaceae may be used in two senses, a wide one (sensu lato) where it refers to legumes as a whole (three subfamilies) or a narrow one (sensu stricto) where it refers to the subfamily Papilionoideae (containing the genus Faba) of the family Papilionaceae. All these legume members carry a variety of traits that necessitate the taxonomic groupings.
These traits include protein content, essential oils content, vitamin composition, medicinal value, ability to fix nitrogen using bacteroids, length of growing season to seed production, cookability, disease resistance /tolerance. The nutrient aspects such protein content, oil content and vitamin content are important traits for human nutrition and thus have been carefully selected for over the past years. A trait such is ability to fix nitrogen has equally been important to agriculturists as this is the trait that allows for soil enrichment and the plant to be used in combination with others as an intercrop. It is important to realise that not all legumes fix nitrogen, those that belong to the Caesalpinioideae sub-family are particularly known not to fix nitrogen, intriguingly so. Cookability traits, use as a source of medicines and length of growing season are the traits that keep changing from geographical location to geographical location since they are influenced by personal choices and other social perceptions. Traits such as resistance or tolerance to disease are important for agronomic production but again this trait is influenced by other environmental biotic and abiotic factors.
Domestication of Tylosema esculentum (marama bean)
Introduction
Marama bean is widely used by the Khoisan and Bantu people of Southern Africa, including Namibia. The plant has a huge potential to address the problem of malnutrition and hunger in Namibia and other dry areas of Southern Africa. Unfortunately marama bean is not yet cultivated but wild, its seeds and tubers are gathered from the wild, although it is important to mention that few isolated activities of cultivating it have stated in Namibia and other places. Some genotypes of the marama are becoming endangered due to the increase in unsustainable harvesting by humans and numbers of range animals that graze on it. Generally, the plant is low yeilding, producing few seeds (one to two seeds per pod). So collecting seed in the wild is not a sustainable way of alleviating the malnutrition problems of this Southern African region. The plant needs to be developed into a crop with cultivars that are high yeilding and early maturing. Molecular characterisation of maramabean using DNA markers seeks to address the problem of identification of the marama bean germplasm in Namibia and beyond. This will be part of a strategy of marama domestication to develop marama as a food crop alternative for dry and sandy areas in the world.
Origin and History
Tylosema esculentum is indigenous to Southern Africa and has been used by the San people for as long as history can tell. It occurs in South Africa (Western, North West and Northern Cape); Botswana (Kgalagadi); Eastern and North-eastern of Namibia (Castro et al, 2005; Amarteifio & Moholo, 1998; Koenen, 2001). The plant grows in sandy soils which have limited water- holding capacity and is frequently exposed to very high light intensity, extreme temperature and prolonged drought (Mitchell et al., 2005). Soils which support growth of marama bean are generally low in organic matter and nutrients, including nitrogen. Marama bean roots do not nodulate and it does not obtain its nitrogen from symbiotic fixation with soil Rhizobia, nor from a source different from that utilized by associated plant species in the same site, but probably scavenges nitrogen efficiently from low concentrations in the soil and rapidly builds reserves in the large tubers to serve as a buffer from formation of protein-rich organs (Dakora et al., 1999). This is particularly intriguing since marama seeds have very high protein content when compared to other nitrogen–fixing plants such as cowpea. How does it manage to harness s nitrogen which such high efficiency?
Biological Descriptions
Tylosema esculentum (Marama bean, also known as gemsbok bean (English); maramaboontjie, elandsboontjie, braaiboontjite (Afrikaans); marama, morama (Tswana); maramama (Thonga), Tsi, tsin (! Kung San); gami (Khoi); ozombanui (Herero) (Van Wyk & Gericke, 2000), is a perennial species, producing a prostrate vine with numerous prostrate stems of up to three meters in length which spread from an enormous woody tuber below the ground (Figure 2) (Powell, 1987). Tubers (Figure 2) have a reddish-brown bark and usually taper to a thinner neck-like structure near the soil surface, from where the annual branches grow during the rainy season. Forked tendrils are found along the stems. The characteristic leaves are deeply two-lobed, hairless, and firm in texture. Attractive bright yellow flowers are born along the stems, each with erect petals and stamens, and are followed by large brown seeds of about 20 millimetres in diameter with a mass of about three grams each. The seed coat is about two millimetres thick, and encloses the delicious and nutritious white nut (Van Wyk & Gericke, 2000). Marama seeds (Figure 2) compete with peanut and soybean in nutritive quality while the young tubers contain protein and are more nutritious than potato or yam. The plant thrives in poor-quality soil and under the harshest of climates (National Research Council, 2006).
 |
Figure 2: A wild patch of Tylosema esculentum (marama bean) with bright yellow flowers (left panel); a root tuber of marama bean (middle panel) and mature brown-coffee marama bean seeds (left panel).
|
Taxonomic position and Habitat
The genus Tylosema (Schweinf.) [Torre & Hillc] comprises of four taxonomically accepted species [T.esculentum (Burch.) A. Schreib., T. fassoglense (Schweinf.) Torre & Hillc., T. argenteum (Chiov.) Brenan and T. humifusum (Pic.Serm. & Roti Mich.) Brenan] occurring in eastern, central tropical and southern Africa (Castro et al., 2005). According to Castro et al. (2005), this genus has only two fertile stamens, the remaining seven or eight stamens being sterile, variously shaped and coloured; with the presence of a lobed non-spathaceous calyx-limb (Castro et al., 2005).
Reproductive Life cycle of Marama bean
In its natural stands, marama bean takes between 18-24 months to reach reproduction maturity. It takes between 8-21 days to germinate on wet soils and then the plant grows vegetatively (Figure 2) for the next 5-6 months. During this time a tuber will be developing underground. This tuber will sustain the plant nutritionally in the coming winter times when all the runners die back and shrivel off. The tuber (Figure 2) will lie in the ground for the next 3-4 months until the next rainy season. At the start of the rainy season the new runners will sprout from the tuber s and marama bean will produce flowers after 1-4 months of vegetative growth. The usually yellow flowers will be pollinated by a solitary carpenter bee and the first pods with seed will set. Thereafter marama bean runners will dieback and re-sprout again as the cycle repeats perennially. The seed usually shatter off a brown dry marama pod powerfully for seed dispersal. The pod starts with a purplish colour, and then it becomes green as the seeds inside mature and final turn brown when the seeds are dispersal-ready.
Karyotype and Genome Size
Marama bean chromosome number and genome size is still subject of research. This crucial basic biological information is required for crop improvement and has not been deciphered. However, it will not be long before it is known. This further underlines the importance of funding basic research for edible plants that have been long neglected as the need to diversify crops is now urgent in the face of fast changing climate and increasing food demands. Furthermore, the genome size of marama bean has not yet determine, pointing to the need for utilising the available deep sequencing platforms that are now available.
Economic Importance of Marama bean
Very limited research has been done on marama including research directed towards domestication, despite its potential as a food and cash crop (Amarteifio & Moholo, 1998). To the tribes in the Kalahari, marama bean contributes 75% of the total vegetable content of the diet. The natives roast the beans in hot sand or coal and the roasted beans eaten at this stage or pounded and boiled with water to make porridge (Vietmeyer, 1986). Seeds (Figure 2) are not usually eaten raw, possibly because of the slimy texture and the disagreeable taste. Strong trypsin inhibition is found in raw seeds but this negative nutritional factor is averted by cooking (Powell, 1987; Bower et al., 1988). The seeds flavour resembles almond or cashew. Some farmers use marama beans as a food supplement for fattening pigs (Elfant et al., 1985). The seeds produces oil with pleasant odour and can be used in food and cosmetic industries (Amarteifio & Moholo, 1998).
The underground tuber of T. esculentum can attain enormous size. Specimens weighing between 150-250 kilograms are common. Young tubers are roasted and eaten (Van Wyk & Gerick, 2000). Livestock and game in Kalahari feed on fresh foliage (National Academy of Sciences, (NAS), 1979) but others state that foliage is not browsed (Powell, 1987). Possibly phenolic compounds in the leaves deter grazing and insects and this may be the reason why people of the Kalahari crush the leaves to make a thick paste that is used to treat wounds and arthritis (Koenen, 2001).
The primary agronomic potential of the marama beans is based upon the high nutritional value of seeds. The protein content of the seed is comparable to or slightly higher than that of soybeans and the oil content of marama beans seeds is twice that of soybeans and approaches that of peanuts (Powell, 1987). The protein content is about 30% to 35% and oil content varies from 35% to 42% (Bower et al., 1988). This underscores the need to domesticate marama bean. Marama beans are a potential source of protein and oil, if available. However, no attempts to systematically cultivate the marama beans have been observed among natives but the beans were sown directly into sand, without ploughing, in the Transvaal, South Africa (Elfant et al., 1985). Graham & Vance (2003) consider that marama bean is a potential crop, presently underutilized, which could be improved and adapted for domestication. Elsewhere in the world, cultivation of marama has been attempted in the arid regions of the USA, Australia and Israel (Monaghan & Halloran, 1996).
Activities related to crop domestication have been transformed by technologies and discoveries in the genome sciences as well as information-related sciences that are providing new tools for bioinformatics and systems biology (Olsen & Schaal, 1999; Paterson et al., 1995; Peloquin, 1981; Casas et al., 2007). Today, an outstanding feature of progress in the sciences related to crop domestication is the availability of well-characterized germplasm resources in the global network of genetic resources centres (gene banks). Germplasm in gene banks is providing research materials for understanding domestication as well as for plant breeding. Progress in this field in recent years has transformed plant breeding to meet the human need for increased crop yield with minimum environmental impacts. It is, however, regrettable that this successful research has neglected plants in Africa, such as marama bean, even though their potential as food and cash crops has been often suggested (Lebutswe et al., 2003; Ramolemana et al., 2003; Nepolo et al., 2009). Usually domestication is a long-term process and this has tended to discourage many workers in this field. However, considering the trend in demand for food and healthy diets, it has become clear to all that domestication to meet needs for food is essential (Piperno &Pearsall, 1998; Pourkheirandish & Komatsuda, 2007; Vaughn et al., 2007).
Domestication and Breeding Objectives for Marama bean
For marama bean, information on its genetic diversity, hybridisation, and speciation in the different population is very limited (Kumerle et al 2008, Nepolo et al., 2009). If any meaningful and time serving plant improvements through breeding and selection can be done to marama, it is critical the the genetic variability of marama is understood first. It has now been finally resolved that there is low interpopulation genetic diversity and high intrapopulation diversity with the genus Tylosema esculentum using microsatellites (Takundwa et al., 2010). Genetic variability, coupled with phenotypic assessements, can be compared in and between populations to gain full insight of the diversity in marama bean. Key traits that can be improved in marams are time to reach reproductive maturity, it is desirable that the plant is able to produce seeds in short time that the current for farmers to maximise benefits from it. However, this must be traded off carefully with retaining all the good characteristics with the seeds. Already, the plant has desirable traits such is surviving under harsh environmental stress of low water (low moisture) and high temperature as well as poor soils.
It is not yet known what is the effeciency of marama bean in utilizing chemical or organic fertilisers. This aspect is important to investigate as it may provide a way of increasing vegetative growth and thus shortening the reproductive cycle. The search for new germplasm that have an early flowering characteistic is yet another avenue of obtaining a variety that mature early. The quality of the seed interms of oil content, protein content, amino acids composition, fatty acid composition has been compared to that of other domesticated legume or even superior in other instances (Bower et al., 1998) (see Table 1).
Table 1: A comparison on legume protein content of marama bean and other major food legumes.
| Legume | % Protein content |
| Soybean (some varieties) | 38 |
| Marama bean | 32 |
| Lupine | 31 |
| Lens | 24 |
| Pea | 23 |
| Broad bean | 23 |
| Phaseolus | 22 |
Germplasm characterization
In the marama bean domestication programme at the University of Namibia, 80 microsatellite primer sets have been developed using modified protocol of Zane et al., 2002 to be used for genetic diversity analysis, germplasm identification and other traits analysis. Over 400 accessions of marama have been collected and microsatellite markers for the characterization of these accession has initiated. In addition, phenotypic characterization of these accessions, phytopathological surveys together with assessment of agronomic performance of the marama accessions has initiated. In the field, phenotype variations in internodal length, number of seeds per pod, number of pods, size of leaves, colour of runners, time of flowering, time to runner die back, leaf green intensity have been observed. However, a study to determine if these variations are genetically stable and inheritable is yet to be done.
Gene Discovery
Partial cDNA sequences of expressed sequence tags of the wild dicotyledonous legume Tylosema esculentum were done using the 454 sequencing platform. This generated over 3000 sequencing including large and small contigs. This is a tremendous resource for mining and discovering new gene for marama bean. Genes of traits of interest, housekeeping regulatory genes can be determined using free internet based search tools such as BLAST. Currently, this is the approach being followed ‘mine’ the database using basic Internet tools to search for genes with specific functions. Already, the identification of the transcription factors and chloroplast specific genes has been done and the search for other genes continues.
Future Scopes of Work
The case for marama bean domestication is clear. The domestication work will continue with the characterization of this plant to understand its growth, development and diversity. The natural diversity at both the phenotypic and genotypic levels will be assessed while simultaneously developing the molecular tools that can be employed in a marker assisted breeding program for the development of cultivars. The phenotypic characterization will focus on yield components such as flower number, seed set, number of seeds per pod and number of seeds per plant. Polymorphic SSRs from marama will be identified and used to develop a molecular map to be a tool for a marker-assisted breeding program aimed at the improvement of this plant.
The SSR markers will be used for a diversity analysis to help in the development of a germplasm collection that represents the maximum diversity of the species in Southern Africa, and to develop a detailed molecular map of marama bean. This map will then be used to develop potential cultivars by combining the best yield attributes (identified through phenotypic screening) using marker-assisted selection.
Concluding remarks
In this article, it has been made clear that the domestication of food legumes has been a long journey for some of the legumes such soybean and cowpea. This has been the so primarily because of lack of tools that could quicken the process. In the case of domestication of marama bean the process is envisaged to be shorter, due to the availability of research tools and the pressure being exerted the ever increasing demand for more food resources. Furthermore, the case of marama bean domestication should be able to serve as an example that there is always need to do research on the little known plants of the world as these may hold the key to solve some of the problems of inhabitants of the harsh environments. However, the availability of funding for such programmes remains a real challenge. One of the broader impacts of the domestication of marama bean will be the availability of a new crop alternative for resource poor farmers in Southern Africa and other arid regions of the world.
Acknowledgments
I would like to acknowledge financial support from the Kirkhouse Trust, United Kingdom for funding the Marama domestication project in Namibia. I would also like to acknowledge Prof Christopher Cullis, Jean-Damascene Uzabakiriho, Dr Martha Kandawa Schulz, Dr David Lawlor, Mutsa Takundwa and Emmanuel Nepolo and Prof Karl Kunert for their support in the domestication project.
References
- Amarteifio JO and Moholo D. 1998. The chemical composition of four legumes consumed in Botswana. Journal of Food Composition and Analysis 11: 329-332.
- Bower N, Hertel, Oh Jand Storey R. 1998. Nutritional analysis of marama bean (Tylosema esculentum), Fabaceae: Analysis of the seed. Economic Botany 42(4): 533-540.
- Casas A, Otero-Arnaiz A, PE´ Rez-Negro ´E N and Valiente-Banuet A. 2007. In situ Management and Domestication of Plants in Mesoamerica. Annals of Botany 100: 1101–1115.
- Castro S, Silveira P, Cuotinho PA and Figueiredo E. 2005. Systematic studies in Tylosem (Leguminosae). Botanical Journal of the Linean Society 147. 99-115.
- Coulibaly S., Pasquet RS, Papa R and Gepts P. 2002. AFLP Analysis of the phenetic organization and genetic diversity of Vigna unguiculata L. Walp. reveals extensive gene flow between wild and domesticated types. Theoretical and Applied Genetics 104: 358-66.
- Dakora FD, Lawlor DW, Sibuga KP. 1999. Assessment of symbiotic nitrogen nutrition in marama bean (Tylosema esculentum L.) a tuber-producing underutilized African grain legume. Symbiosis 27: 269–277.
- Elfant, M., Bryant. L, and Starcher, B. 1985. Isolation and characterization of protease inhibitors of the Marama bean. Tylosema esculentum). Proceedings of the Society for Experimental Biology and Medicine 180(2):329-333.
- Gepts P. 2004. Crop Domestication as a Long-term Selection Experiment. Plant Breeding Reviews, Volume 24, Part 2.
- Graham PH and Vance CP. 2003. Legumes: Importance and Constraints to Greater Use. Plant Physiology Vol. 131, pp. 872–877.
- Hymowitz T. 1970. On the domestication of soybean. Economic Botany Vol. 24, No. 4, pp. 408-421
- Koenen, E. V. (2001). Medicinal poisonous and edible plants in Namibia. Klaus Hess Publisher/ Verlag Windhoek- Gottingen.
- Kumerle B, Carrino S, Chimwamurombe PM & Cullis CA. 2008. Developing DNA markers for underserved crops important for the developing world. (http://ora.ra.cwru.edu/showcase/showcasedetail.asp?type=post&id=1312).
- Lebutswe T., Ramolemana G.M., Machacha S. and Tsopito C.M. 2003. Effects of breaking dormany treatments on the germination and emergence of morama bean (Tylosema esculentum burch). UNISWA Journal of Agriculture 11: 28-33.
- Mitchell, R.A.C.; Keys, A.J.; Madgwick, P.J.; Parry, M.A.J.; Lawlor, D.W. 2005. Adaptation of photosynthesis in marama bean Tylosema esculentum (Burchell A. Schreiber) to a high temperature, high radiation, drought-prone environment . Plant Physiology and Biochemistry 43 (10-11) pp. 969-976.
- Monaghan BG and Halloran GM. 1996. RAPD variation within and between natural populations of morama (Tylosema esculentum[Burchell]Screiber) in Southern Africa. South Africa Journal of Botany 62(6): 287-291.
- National Academy of Sciences. 1979. Tropical legumes: Resources for the future. A report of an Ad Hoc Panel of the Advisory Committee on Technology for International Development Commission on International Relations, National Council Research, Library of Congress Catalogue Number 79-64185.
- National Research Council. 2006. Lost Crops of Africa: Volume II: Vegetables. National Academies Press, Washington, DC. Chapter 13, Marama pp 234-245
- Nepolo E, Takundwa M, Chimwamurombe PM, Cullis CA and Kunert K. 2009.. A review of geographical distribution of Marama bean [Tylosema esculentum (Burchell) Schreiber] and genetic diversity in the Namibian germplasm. African Journal of Biotechnology Vol. 8 (10), pp. 2088-2093.
- Olsen KM., and Schaal B. 1999. Evidence on the origin of cassava: phylogeography of Manihot esculenta. Proc. Natl. Acad. Sci. (USA) 96:5586-5591.
- Paterson AH., Lin YR, Li ZK, and Schertz KF. 1995. Convergent domestication of cereal crops by independent mutations at corresponding genetic loci. Science 269:1714-1718.
- Peloquin, SJ. 1981. Chromosomal and cytoplasmic manipulations. p. 117-137. In: K. Frey (ed.), Plant breeding II. Iowa State Univ. Press, Ames.
- Piperno, D., and Pearsall D. 1998. The origin of agriculture in the Neotropics. Academic Press, San Diego.
- Pourkheirandish M and Komatsuda T. 2007. The Importance of Barley Genetics and Domestication in a Global Perspective. Annals of Botany 100: 999–1008.
- Powell AM. 1987. Marama bean (Tylosema esculentum, Fabaceae): Seed crop crop in Texas. Economic Botany 41: 216-220.
- Ramolemana G., Machacha S., Lebutwse T. and Tsopito C.M. 2003. Response of marama bean (Tylosema esculentum) to phosporus and nitrogen utilisation in a sandy loam at Sebele, Botswana. UNISWA Journal of Agriculture 11: 34-39.
- Takundwa, M., Chimwamurombe, P.M., Kunert, K. & Cullis, C.A. 2010. Isolation and characterization of microsatellite repeats in Marama bean (Tylosema esculentum). African Journal of Agricultural Research. Vol. 5(7) pp. 561-566.
- Vaughan DA, Bala´ ZS and Heslop-Harisson JS. 2007. From Crop Domestication to Super-domestication. Annals of Botany 100: 893–901.
- Van Wyk, B., and Gericke, N. 2000. People’s Plants: A guide to useful plants of southern Africa. Briza publications, South Africa. pp. 26.
- Vietmeyer, N. D. 1986. Lesser- Known Plants of Potential use in Agriculture and Forestry: Science, Vol. 232:1379-1384 article 1379.
- Zane L., Bargelloni L. and Patarnello T. 2002. Strategies for microsatellite isolation: a review. Molecular Ecology 11: 1-16.

This work is licensed under a Creative Commons Attribution 4.0 International License.



