

Manuscript accepted on : 04 December 2017
Published online on: --
Apurva Sharma , Seema Langer and Nipoon Sharma
, Seema Langer and Nipoon Sharma
Department of Zoology, University of Jammu, 180006.
Corresponding Author E-mail: apurvasharmajmu@gmail.com
DOI : http://dx.doi.org/10.13005/bbra/2595

ABSTRACT: An investigation has been conducted on the Gho-Manhasan stream of Jammu region from Dec 2014- Nov 2015 with an aim to work out the various breeding related changes in the Himalayan prawn Macrobrachium dayanum. During this period, certain important periodical modifications were found likely as the presence of extra setae especially for the purpose of egg bearing. The special breeding dress came into being after the pre-breeding moult. The eight groups of breeding setae have been observed as Simple setae, Pappose setae, Spiniform setae, Protective setae, Plumose setae, Serrate setae, Multidenticulate setae and Ovigerous setae.
KEYWORDS: Macrobrachium Dayanum; Breeding Dress; Setae
Download this article as:| Copy the following to cite this article: Sharma A, Langer S, Sharma N. Presence of Various Type of Setae as A Periodical Modification in Himalayan Prawn Macrobrachium Dayanum from Jammu Region. Biosci Biotech Res Asia 2017;14(4). |
| Copy the following to cite this URL: Sharma A, Langer S, Sharma N. Presence of Various Type of Setae as A Periodical Modification in Himalayan Prawn Macrobrachium Dayanum from Jammu Region. Biosci Biotech Res Asia 2017;14(4). Available from: https://www.biotech-asia.org/?p=28592 |
Introduction
Many decapod crustacean species spend considerable time engaged in grooming or cleaning. These species have a variety of structures and behaviors that function to prevent body fouling by epizoites and particulate debris. During grooming, the decapod scrapes its exoskeleton with brushes of different types of setae appears at certain period of time, mostly during breeding season. During the breeding period the integument of the female Macrobrachium dayanum went important modifications characterized by the presence of extra setae on different body parts. Some of the setae appear for the purpose of egg bearing. This special breeding dress came into being after the pre- breeding moult preceeding copulation and ovulation while, some setae disappear after the breeding is over. Such a breeding dress is found not only in Palaemonetes but also in other Palaemonidae and throughout Caridea (Yonge, 1955). These setae were observed to be periodical in nature and appears only after pre breeding moult as they were not among permanent secondary sexual character. According to Watling (1989), a seta is ‘an articulated with a basal socket cuticular extension of virtually any shape or size’ whereas a spine always lacks a basal socket. By this definition many of the spines, spinules, tubercles and scales described by previous authors on the second chelipeds and body of Macrobrachium are clearly setae. The integument of the female Macrobrachium dayanum also undergoes important modifications pertaining to the presence of extra setae.
Material and Methods
The live specimens of the prawn Macrobrachium dayanum were collected from the Gho-Manhasan stream of Jammu region (J&K). The collected specimens were immediately preserved in 10% ethyl alcohol. The first to fifth pleopods were removed from the body and the temporary slides were made. The slides were then scanned and micro photographed with Sony SSC-DC 378P digital camera attached with microscope provided by the department.
Results and Discussion
Several systems for the classification of setae have been proposed, some of the recent ones include those of Thomas (1970), Watling (1989) and Felgenhauer (1992). The following classification of seta in Macrobrachium is largely based on the setal types diagnosed by Thomas (1970) and Felgenhauer (1992). During the present investigation it was observed that the integument of the female Macrobrachium dayanum went important modifications characterized by the presence of extra setae for the purpose of egg bearing. The special breeding dress came into being after the pre- breeding moult. These setae were observed to be periodical in nature and appear only after pre breeding moult as they were not among permanent secondary sexual character. Eight groups of breeding setae have been observed which are as follows as shown in figures.
Simple Setae
Simple setae (Fig 1) have smooth setal shafts without setules or scales and taper to a sharp point. In Macrobrachium dayanum several smooth setae were observed on the periopods. The most ubiquitous type of smooth seta was long and moderately stiff with a sharp point like a typical bristle.
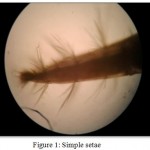 |
Figure 1: Simple setae
|
Pappose Setae
Pappose setae (Fig 2) are generally long, fine and thread-like. In Macrobrachium dayanum they typically occur together in dense clumps on the segments of the second pereiopod. The shaft of pappose setae bears numerous long, randomly arranged setules.
 |
Figure 2: Pappose setae
|
Spiniform Setae
These are a robust form of smooth seta (Fig 3) which is spine-like in appearance. In Macrobrachium dayanum these setae were evident as they usually occur in low numbers at fixed positions on the dorsal and posterior telson. Previous authors have generally referred to spiniform setae as spines or spinules.
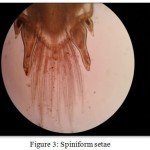 |
Figure 3: Spiniform setae
|
Protective Setae
Protective setae (Fig 4) were also observed in Macrobrachium dayanum. The stout smooth setae were abundant on the segments of the second pereiopod in developed male. Protective setae show considerable variation in size and shape. Generally they first appear as small spinule like buds but increase in relative size as the chelipeds develop. In developed males of many species these setae are also found on the anterolateral carapace and abdomen. Unlike spiniform setae, which tend to occur in fixed positions in low numbers, these setae are found in large numbers. Protective setae are termed as they appear to have a defensive function against predators, similar to true spines and teeth. Protective setae are always heavily tanned and lack an annulus or basal septum. They have generally been referred to as spines, spinules, tubercles or scales by previous authors.
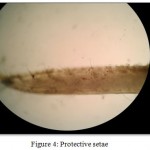 |
Figure 4: Protective setae
|
Plumose Setae
It was the most common type of compound seta in this prawn. Plumose setae (Fig 5) bear two distinct rows of long, usually delicate setules whose insertions are always directly opposite one another on the setal shaft. The shaft is typically devoid of setules towards the base. Plumose setae or ‘feathered’ setae are commonly found along the margins of natatory appendages such as the pleopods in Macrobrachium dayanum. Plumose setae appear to improve swimming efficiency by extending the effective surface area. The setal shaft is robust and spine-like, with the setules arranged in two rows set at an angle of approximately 90 to each other. The setules are absent near the tip and on the basal third of the setal shaft.
 |
Figure 5: Plumose setae
|
Serrate Setae
This is another very common type of compound seta characterised by the presence of sharp, relatively short, denticules arising from the setal shaft and a basal septum. The denticules always arise distal to a well-developed annulation half- way along the shaft .Serrate setae (Fig 6) are commonly found on appendages important in grooming the exoskeleton. Typical serrate setae have serrations arranged in two opposite rows, set at approximately 45 to one another, gradually falling to the horizontal distally. These setae typically occur on the terminal segment of the third maxilliped and proximal propodus and distal carpus in Macrobrachium dayanum.
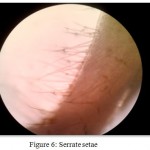 |
Figure 6: Serrate setae
|
Multidenticulate Setae
Fig 7: These types of compound setae were present on the minor second pereiopods of the investigating species. Typically, they were restricted to the cutting edges of the fingers, but in some cases are also found elsewhere on the minor cheliped. Unlike serrate setae, the denticules or scales are placed around the shaft in a random or spiral pattern.
 |
Figure 7: Multidenticulate setae
|
Ovigerous Setae
When the Macrobrachium dayanum is ready to spawn, it develops a breeding dress which is the most outstanding feature that results in the emergence of a set of ovigerous setae (Fig 8) on pleopods usually on pleopod I, pleopod II and Pleopod III at the time of the pre- spawning moult. At the time of spawning the eggs are attached to these setae which hold them until hatching (Fig 9). Ovigerous setae only appear when the female moults into egg carrying conditions (Fig 10). After pre spawning moult ovigerous setae are developed on the inner margins of the bases of the first three pairs of pleopod which serve to anchor the eggs to the pleopod during incubation. It appears that eggs which are sticky adhere to each other and to ovigerous setae upon entering the brood chamber. The basal segment of first three pairs of pleopod get elongated and are provided with tufts of long ovigerous setae on their inner margins and the egg mass is attached to these branched setae.
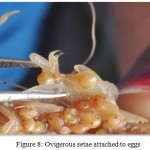 |
Figure 8: Ovigerous setae attached to eggs
|
 |
Figure 9: Curling of setae for the attachment to eggs
|
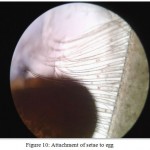 |
Figure 10: Attachment of setae to egg
|
Our results get support from various other workers viz. Nagamine and knight, (1980) and Belanger et al. (2008) who also have authenticated different setae particularly ovigerous setae as the breeding dress components in various prawn species. Nagamine and Knight (1980) made a detailed study on sexually dimorphic structures of the Malaysian Prawn, Macrobrachium rosenbergii and concluded that the brood chamber and reproductive setae of females begin to develop at 20 mm CL which is recorded at several molts before their first spawning. He divided reproductive setae into ovigerous and ovipositing setae, depending upon their function. Present author also hold the viewpoint that ovigerous setae are temporary structures formed only during preparturial molts, some ovipositing setae are also temporary while, others are permanent since they are retained on nonparturial molts. Variations however in their length are apparent at different stages of development.
Present observations also get support from those recorded by Thomas (1970) who showed that smooth setae are widely distributed, but are particularly well developed on the distal corners of limb segments. Hindley and Alexander (1978) found that distally, the groups of smooth (simple) setae on the chelae of the banana prawn (Penaeus merguiensis) lie close together and contain up to 15 setae. They also found that the distribution of the groups became sparser proximally. Presently in Macrobrachium dayanum, it was seen that distal portion of setae were equipped with dense bunches of predominantly smooth setae, while proximal portion contained considerably fewer smooth setae, evenly mixed with plumose setae. Plumose setae are found evenly distributed across the surface of the major chelae in males. Plumose setae contain setules which function to increase the surface area of sensory appendages. Present observations regarding the smooth and plumose setae reveal that both op. cit groups of setae are densely present over natatory appendages i.e pleopods. Belanger et al. (2008) anlaysed that the major chelae of reproductive male crayfish contain both smooth and plumose setae and are used for the perception of female odors in crayfish, Orconectes rusticus. Further it was observed that there is a significant difference in the number and distribution of sensory setae types among reproductive forms, but the types of sensory setae found on the chelae of both forms are similar.
Present author also holds the view point that the tip of each seta has an elaborate structure equipped with a large sub-terminal pore which enables them to perform chemosensory function responsible for discriminating edible from non-edible material. Similar viewpoint has been witnessed earlier by Hindley and Alexander (1978) who have reported three groups of differently sculptured setae, at the articulation of the propodus and carpus of the first pereiopod whose function is to clean the other chelate pereiopods. Crustaceans are covered with rigid exoskeletons and receive external sensory input through cuticular hair organs referred to as setae, located on sensory appendages. In M. dayanum, in particular, the major chelae have mechano and chemosensitive setae and may contain cuspidate (also referred to as toothed, squat, or fringed setae), hemate, smooth and plumose setae as has also been reported by Belanger and Moore (2006) and Altner et al. (1983). According to Watling (1989) the setae are found over the entire surface of crustaceans, including the major chelae, and serve a wide range of mechano- and chemosensory functions. Most studies on the major chelae of crayfish have focused on their mechanical function during reproduction (Keller and Hazlett, 1996), although a recent study has shown that the major chelae are used to detect and discriminate female odors in reproductive (form I) male crayfish. Non-reproductive (form II) males do not show this ability Form I males have more pockets of sensory setae on their major chelae than form II males. In M. dayanum also it has been observed that when a male moults into the reproductive form the number of sensory setae on the chelae increase considerably. All these findings suggest that the major chelae are important structures for mating (Keller and Hazlett, 1996; Belanger and Moore, 2006). Hazlett (1975) described that the major chelae have an important function in the perception of female odors during reproduction and may provide additional chemical feedback necessary for discrimination of female stimuli.
References
- Altner H, Hatt H and Altner I. Structural and functional properties of the mechanoreceptors and chemoreceptors in the anterior esophageal sensilla of the crayfish, Astacus astacus. Cell and Tissue Research. 1983;228:357-374.
- Belanger R. M and Moore P.A. The use of the major chelae by reproductive male crayfish (Orconectes rusticus) for discrimination of female odours. Behaviour. 2006;143:713-731.
CrossRef - Belanger R. M, Ren X, Mcdowell K, Chang S, Moore P and Zielinski B. Sensory setae on the major chelae of male crayfish, Orconectus rusticus (Decapoda : Astacidae). Journal of Crustacean biology. 2008;28(1):27-36.
CrossRef - Felgenhauer B.E. External anatomy and integumental structures in Decapod Crustacea, Microscopic Anatomy of Invertebrates. New York (Wiley-Liss). 1992;10:7-43.
- Hazlett B.A. Ethological analyses of reproductive behavior in marine Crustacea. Pubblicazioni della Stazione Zoologica di Napoli. 1975;39 (Supplement):677-695.
- Hindley J.P.R and Alexander C.G. Structure and function of the chelae periopods of the banana prawn. Penaeus merguiensi Marine Biology. 1978;48:153-160.
CrossRef - Keller T. A and Hazlett B.A. Mechanical use of crayfish chelae. Marine and Freshwater Behaviour and Physiology.1996;28:149-162.
CrossRef - Nagamine C.M and Knight A.W. Development, Maturation and function of some sexually dimorphic structures of the Malaysian Prawn, Macrobrachium rosenbergii (De Man) (Decapoda, Palaemonidae). Crustaceana. 1980;39(2):141-152.
CrossRef - Thomas W. J. The setae of Austropotamobius pallipes (Crustacea: Astacidae). Journal of Zoology.London. 1970;160:91-142.
CrossRef - Watling L. A classification system for crustacean setae based on the homology concept, Functional morphology of feeding and grooming in Crustacea. A.A Balkema Publisher, Rotterdam. 1989;15-26.
- Yonge C.M. Egg attachment in Cranqon vulgaris and other Caridea. Proceedings of the Royal Society of Edinburgh. Section B (Biology). 1955;65, pt .3(24):369-400.

This work is licensed under a Creative Commons Attribution 4.0 International License.



